





文摘
beryllium-exposed工人发展铍敏化作用的一个子集(BeS)先于慢性铍病(CBD)。我们进行了一次深入分析CBD的候选基因的异常表达。
我们进行了Affymetrix GeneChip第1.0阵列分析外周血单核细胞(PBMCs)从10 CBD, 10 BeS和10 beryllium-exposed, nondiseased控制与BeSO刺激4或中等。差异表达基因的高通量实时PCR验证这一组额外的情况下和nonexposed控制。CBD最大的候选基因的功能角色使用药理抑制剂进行了评估。CBD基因表达数据与全血和肺组织结节病的基因表达综合。
我们证实了几乎450个基因之间的明显差异表达CBD和控制。基因是木菠萝的最高浓缩(Janus激酶)统计(转录信号传感器和催化剂)信号。JAK2抑制剂显著降低肿瘤坏死factor-α和interferon-γ生产。此外,我们发现287个差异表达基因重叠在CBD /结节病。顶部共享途径包括cytokine-cytokine受体相互作用,和toll样受体,趋化因子和JAK-STAT信号通路。
我们表明,PBMCs演示相关差异表达基因档案immunnopathogenesis CBD。CBD和结节病分享相似的致病基因和通路的微分表达式。
文摘
慢性铍病和结节病相似致病基因和通路的微分表达式http://ow.ly/YgjIA
介绍
慢性铍病(CBD)、肉芽肿性肺部疾病引起的铍暴露在工作场所,特点是beryllium-specific CD4的积累+t细胞在肺。根据遗传易感性和曝光,CBD发生在多达20%的暴露工人(1]。铍敏化作用(BeS)先于CBD的特点是积极铍淋巴细胞增殖试验(BeLPT) [2]。BeS发展作为铍的外围的免疫反应,但没有证据表明肺部疾病和/或肉芽肿在肺活检。喜神贝斯的流行范围从2到20%的暴露工人(2- - - - - -4]。BeS进展CBD每年大约8%的速度1(5]。
一旦CBD CD4+肺被激活t细胞,这些细胞无性繁殖系地增殖和分泌辅助1 (Th1)类型细胞因子白介素(IL) 2、interferon-γ和肿瘤坏死因子(TNF) -α[6- - - - - -8),导致巨噬细胞激活,积累、聚合和肉芽肿性炎症的发展9]。铍暴露刺激多种细胞反应,包括细胞迁移(10),细胞因子调节(11)和生长抑制(12]。这些免疫调节活动支持了这样的观点,即触发铍non-CBD个人的先天免疫反应以及获得性免疫反应。
其他基因的功能和表达的调控在喜神贝斯和CBD是未知的,只有有限的beryllium-related研究已经进行到目前为止(13,14]。全基因组表达模式研究了在结节病,一种原因不明的疾病,表现为noncaseating肉芽肿,主要在肺部(15- - - - - -18]。CBD和结节病的临床、放射学、组织学相似之处,我们提出CBD会共同致病的途径与结节病和特定疾病模式。
我们的目标是调查CBD外周血单核细胞的基因表达谱(PBMCs)来定义相关基因和模式CBD与结节病重叠,这将提供洞察疾病发病机理和协助临床表现型。
材料和方法
研究人口和设计
所有参与者给通知书面同意依照赫尔辛基宣言和这项研究是国家犹太卫生机构审查委员会批准人类受试者(HS # 2435)。20 CBD患者,20 BeS病人和20个对照组(Con),包括10 beryllium-exposed nondiseased控制(ND-Con beryllium-exposed控制没有CBD或喜神贝斯)和10 nonexposed nondiseased控制(NE-Con nonexposed控制没有CBD或喜神贝斯),参加本研究(图1)。试图频率配合控制和用例年龄和性别。
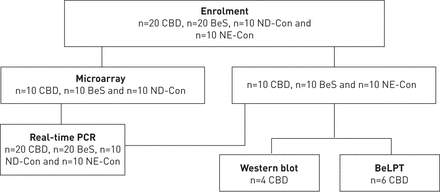
流程图和研究受试者参与研究的利用率。生物多样性公约;慢性铍病;贝斯:铍敏化作用;ND-Con: beryllium-exposed nondiseased控件;NE-Con: nonexposed nondiseased控件;BeLPT:铍淋巴细胞增殖试验。
两阶段研究是设计与每组10科目参加第一阶段。这些记录与CBD与喜神贝斯和控制验证在第二批20每组受试者,包括最初的10每组每组和10个新科目,科目使用高通量实时PCR。
方法
疾病诊断和排除
CBD的诊断成立使用之前定义的标准,包括铍暴露史、肉芽肿性炎症的存在肺活检,和积极的增殖反应的血液和/或支气管肺泡灌洗(BAL)细胞BeSO4在体外。喜神贝斯的诊断建立了基于历史的曝光,铍BeSO PBMCs的积极增生性反应4在体外BeLPT和缺乏肉芽肿性炎症或其他异常的肺活检。我们排除了主题与系统性免疫抑制剂治疗CBD,因为这可能会影响基因的表达。一些人用吸入糖皮质激素治疗。我们排除了受试者如果他们使用烟草(吸烟或其他)在过去的6个月。
PBMC收集和铍的治疗
血液样本被吸引和PBMCs隔绝heparinised Ficoll-Hypaque分离血液。PBMCs (1×106毫升1与100年)刺激μM BeSO4单独或中等和培养24小时(19]。
RNA制备
的总RNA提取PBMCs RNeasy迷你工具根据制造商的协议(试剂盒)。
探针制备、杂交
探针制备、杂交进行基因组学和微阵列的核心在科罗拉多大学丹佛使用GeneChip第1.0数组(Affymetrix、威、英国)。
高通量实时聚合酶链反应
进行高通量实时PCR验证候选基因使用TaqMan™OpenArray©NT循环系统(美国应用生物系统公司,培育城市,CA) (20.]。
西方墨点法
PBMCs暴露在100年μM BeSO4为0,1,2,6 - 24 h,然后收获试验(21]。
铍淋巴细胞增殖试验
细胞在没有孵化或存在1μM TG101348 30分钟和治疗无民事行为能力人或者10或100μM BeSO44天。其他地方扩散测量所述(19]。
统计分析
数据(除了微阵列实验)由单向方差分析进行分析,其次是菸害的测试事后比较。统计分析使用商用软件(Statview v5.0.1;美国NC SAS研究所,卡里)。假定值< 0.05为显著和测试都是双面的。
微阵列分析
的强度探测芯片样本日志2转换,表达式正常化和估计使用健壮的multiarray分析方法应用于样品调整实验使用R版本2.13.0可变性(https://cran.r-project.org)。微阵列样本按CBD-stimulated分组/ CBD-unstimulated BeS-stimulated / BeS-unstimulated和Con-stimulated / Con-unstimulated。我们为多个比较使用Benjamini-Hochberg过程调整。对于每一个结果,我们首先rank-ordered根据假定值的差异表达基因,然后评估这些基因调节或下调> 1.5倍以下的分析表达强度(22- - - - - -24]。每一元的名义显著性水平测试是p≤0.05和置信水平的错误发现率(罗斯福)为90%。
血液比较转录组与结节病
每个比较基于基因表达水平调整假定值比较差异表达的成绩单从结节病与控制从K另外一些et al。(17),从基因表达综合下载(GEO)。(http://www.ncbi.nlm.nih.gov/sites/GDSbrowser?acc=GDS4587)。例活检证实。
比较肺组织转录组与结节病
每个比较基于假定值调整的表达水平与386年相比差异表达转录本在分析由C激励者et al。(18),从地理下载(http://www.ncbi.nlm.nih.gov/sites/GDSbrowser?acc=GDS3580)。例活检证实。
基因集富集分析所描述的其他地方(25,26使用基于网络的软件)(http://www.broad.mit.edu/gsea/)。
功能分类和路径分析是评估基因本体论(去)和《京都议定书》百科全书的基因和基因组(KEGG)通路分析数据库中的注释,可视化和综合发现(大卫)生物信息学资源(http://david.abcc.ncifcrf.gov)[27]。
路径分析和网络一代重叠基因与创造力的途径在CBD和结节病进行分析软件(http://www.ingenuity.com)。
结果
病人的人口统计
受试者所示的人口表1。没有区别是喜神贝斯的年龄和CBD病人参加本研究。大多数学科组都是男性,典型的铍工业。无显著差异的估计之间的持续时间从第一观察铍暴露BeS和CBD科目。CBD受试者的百分比显著增加BAL患者淋巴细胞而喜神贝斯(17.9±13.0与7.6±5.9,p < 0.01)。结节病的临床特点是S1a指出在线补充表。平均肺功能相似CBD与结节病病例。结节病病例从K另外一些et al。(17)和低肺功能正常,在系统性治疗和强的松(在线补充表S1a)。结节病病例K另外一些et al。(17)和CBD吸入型皮质类固醇激素的情况下显示出类似的用法。
基因表达谱证明著名CBD与喜神贝斯相比,反应和喜神贝斯和控制PBMCs之间的相似之处
我们分析三组中PBMCs (CBD-stimulated, BeS-stimulated和Con-stimulated)显示472个重要基因(罗斯福< 10%,p < 0.05)(在线补充表S1)。比较CBD-stimulated与BeS-stimulated PBMCs透露797年重要的基因,其中包括661调节和136个基因表达下调。只有47个基因差异表达> 1.5倍,与42调节和五个表达下调(在线补充表印地和c)。我们观察到735 CBD-stimulated之间明显的差异表达基因和Con-stimulated PBMCs。有趣的是,543个基因调节和191个表达下调,237个差异表达> 1.5倍(233年调节和四个表达下调的基因;在线补充表S2a和b)。CBD-stimulated PBMCs证实细胞因子/趋化因子基因的表达水平显著高于Con-stimulated PBMCs,包括CXCL10,CXCL9,CCL8,CCL3,肿瘤坏死因子,STAT2和SOCS3。只有三个细胞因子/趋化因子基因(CCL3,CCL3L1和SOCS1)在CBD-stimulated更高与BeS-stimulated PBMCs。
37岁的比较BeS-stimulated和Con-stimulated PBMCs基因差异表达> 1.5倍,18日调节和19个表达下调基因(数据未显示)。CBD-stimulated和BeS-stimulated PBMCs演示了一个Th1和增加免疫反应STAT1,肿瘤坏死因子,IRF1,IRF4,IRF8和ICAM1表达式。图2显示了CBD-stimulated基于KEGG通路基因表达模式与Con-stimulated和BeS-stimulated PBMCs(罗斯福< 5%,p < 0.001)。CBD-stimulated基因表达是明显不同于BeS-stimulated Con-stimulated基因表达,Con-stimulated BeS-stimulated最相似。在我们的研究中,我们没有发现IL-17通路的激活,这证实了我们的t细胞immunophenotype研究CBD(数据进行审查)。我们有特征的表达从患者的肺部和外周血Th17商务区在我们的研究中。IL-17没有显著增加+CD4+t细胞检测到在CBD的记忆t细胞,这是符合我们当前的基因表达数据。
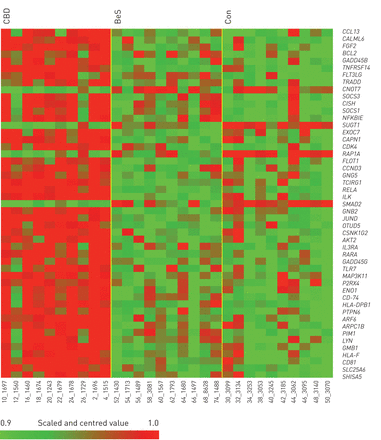
代表表达的热图beryllium-stimulated铍敏化作用(BeS),慢性铍病(CBD)和控制(Con)外周血单核细胞由微阵列分析。红色代表表达增加;绿色表示nonfailing总体均值的表达降低。梯度表明基因表达水平随时间变化的。数据在阈值范围:-2.7 - 2.9。
比较的CBD-unstimulated BeS-unstimulated PBMCs,只有9基因(三个调节和6个表达下调)差异表达> 1.5倍(在线补充表S3a和b)。有55个基因(调节和28表达下调27日)差异表达CBD-unstimulated > 1.5倍与Con-unstimulated PBMCs,而没有差异表达基因在BeS-unstimulated > 1.5倍与Con-unstimulated PBMCs(在线补充表S4a和b)。
基因集的分析和功能分析显示新的和以前在CBD-stimulated确定候选基因与Con-stimulated PBMCs
基于以上研究结果,161功能基因集,代表117个分子功能或766生物过程,比较CBD-stimulated和Con-stimulated PBMCs。分析显示,这些基因主要参与功能leukocyte-mediated细胞毒性或生物过程相关,T-cell-mediated细胞毒性,细胞死亡,cytokine-mediated信号通路和调控和抗原/蛋白质/翻译因素活动核酸/主要组织相容性复合体(MHC)蛋白质绑定(图3)。使用大卫CBD-stimulated Con-stimulated表达水平进行比较,我们发现更多的免疫系统流程、免疫反应、防御反应,反应压力、炎症反应和应对外部刺激,伤害其他生物,病毒和细菌(p < 0.0001)。18丰富KEGG的最重要的途径是木菠萝(Janus激酶)统计(转录信号传感器和催化剂)通路,包含15个基因(p = 0.000003;表2)。
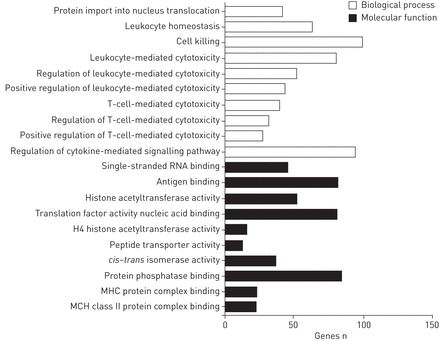
基因本体论范畴分析相关候选基因发现beryllium-stimulated慢性铍病和对照组之间的差异。MHC:主要组织相容性复合体。这些功能基因集被确定的161位宾客代表共有117个分子功能或766生物过程。排名前十的生物过程和前十的分子功能。
CBD和结节病转录概况、通路和网络
差异表达基因在CBD和结节病被Entrez基因数量相比,287个差异表达基因CBD和结节病PBMCs之间共享,包括271调节和16个基因表达下调(在线补充表S5a和b)。监管机构NFKBIE,ICAM1和LILRB4,而顶部通路cytokine-cytokine受体相互作用,和toll样受体,趋化因子和JAK-STAT信号通路。有14个基因调控方向相反(在线补充表S5c和d)。评估这些差异表达基因/通路在CBD和结节病显示重叠网络,参与炎症反应,免疫反应,脂质代谢和分子运输。顶部重叠网络(图4)包括统计,CISH,SOCS1,SOCS2,SOCS3,肿瘤坏死因子和其他人。STAT转录因子作为中心分子连接。
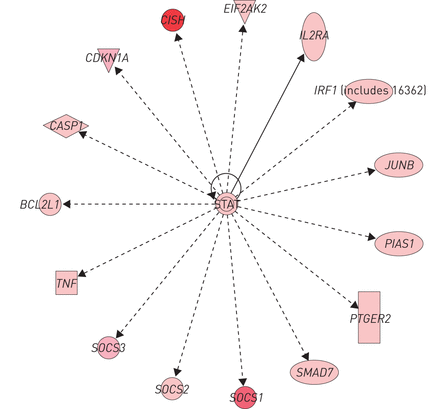
独创性顶部重叠网络的通路分析慢性铍病和结节病。以图形方式显示网络节点(基因/基因产物)和边(生物节点)之间的关系。节点的颜色强度表明褶皱(红色代表upregulation)基因的表达变化。
此外,我们研究了从结节病肺组织的差异表达基因在我们的CBD概要文件。我们发现33个差异表达基因重叠在CBD PBMCs和肺结节病,其中包括13个(包括调节基因CXCL9,FAM26F,TAP1,CXCL11,4英镑,CCL8和STAT1)和一个表达下调的基因。有19个基因调控方向相反(在线补充表S5e-h)。
最高的候选基因的异常表达在一组独立的验证CBD,喜神贝斯和对照组
我们证实了近450的526个基因,我们发现上面使用实时PCR。最高的基因包括褶皱变化CXCL9,HCAR3,HCAR2,CD274,CLEC6A和EGR2CBD-stimulated在统计学上显著性差异和Con-stimulated组(图5)。一些样品有相似的价值观,导致“骗”的重叠的符号组(如。图5 a)。如上所述,BeS受试者没有统计学上不同于CBD或控制,但显示比控制变量的响应。
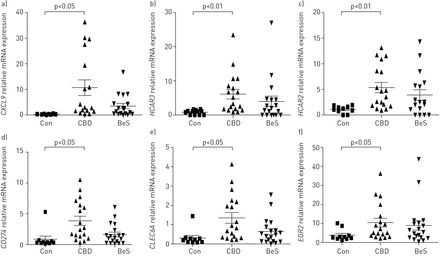
验证的慢性铍病(CBD)候选基因使用OpenArray系统:a)CXCL9,b)HCAR3c)HCAR2d)CD274,e)CLEC6A和f)EGR2。反对:control-stimulated组;CBD: CBD-stimulated组;贝斯:铍sensitisation-stimulated组。这些前六基因过表达在CBD显示为代表我们的发现为我们验证。每个符号代表一个单独的主题;酒吧展示意味着±扫描电镜。
JAK2-STAT1通路是CBD的功能
评估的功能作用通路和基因,我们评估四CBD PBMCs JAK2蛋白质磷酸化蛋白免疫印迹。磷(p)的表达jak2 / p-STAT1增加1 h铍刺激后,虽然SOCS3增加后6 h (图6)。JAK2抑制剂TG101348显著降低了扩散PBMCs 10和100年后的BeLPTμM铍治疗(n = 6),这表明木菠萝通路JAK2基因导致持续的炎症在应对铍肺(图6 b)。
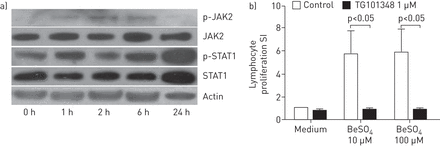
)Janus激酶(激酶)2 /信号传感器和激活转录(STAT) 1磷酸化是铍在外周血单核细胞(PBMCs)慢性铍病(CBD)的病人。PBMCs处理100μM BeSO4表示时间和细胞溶解产物含有等量的蛋白质被使用抗体免疫印迹分析磷(p) JAK2、总JAK2 p-STAT1 STAT1和肌动蛋白。总JAK2和STAT1作为加载控制。b) JAK2抑制剂TG101348下调在CBD (n = 6)淋巴细胞增殖。PBMCs没有孵化(“介质”)或存在1μM TG101348 30分钟,并没有处理或10和100年μM BeSO44天。数据意味着±扫描电镜刺激指数(SI)。
讨论
在这项研究中,我们表明,喜神贝斯和CBD演示Th1基因组转录概况,上级与细胞因子和趋化因子在CBD比控制。具体来说,CBD中差异表达的基因主要是参与相关功能或生物过程leukocyte-mediated细胞毒性,T-cell-mediated细胞毒性,细胞死亡,cytokine-mediated信号通路及其监管。最重要的途径是JAK-STAT通路,是重要的细胞因子调节。毫不奇怪,顶部与CBD相关的基因包括细胞因子/趋化因子和信号的基因,如CXCL10,CXCL9,肿瘤坏死因子,STAT1,STAT2和SOCS3。此外,这些基因/通路是明显的在另一个肉芽肿性肺部疾病,即。结节病,几乎300个基因在两种疾病认识提高监管。最后,我们证实JAK2的功能方面,证明改变这些基因影响beryllium-stimulated细胞因子生产和细胞增殖。关于CBD的发病机制提供重要的数据信息,与类似的发现在结节病,影响其他exposure-mediated免疫疾病。
我们确定了浓缩的基因参与JAK-STAT, cytokine-cytokine受体相互作用,抗原处理和表示,toll样受体,并点头(nucleotide-binding oligomerisation域)——受体信号通路以及基因和通路而不是以前被认为与CBD,等SOCS3和STAT1。JAK-STAT通路的异常激活与自身免疫和炎症性疾病,如风湿性关节炎(28,29日,系统性红斑狼疮30.和克罗恩氏病31日),和其他肺肉芽肿性疾病,包括肺结核(32和结节病32,33]。此外,JAK2抑制剂TG101348减少beryllium-induced扩散在CBD,木菠萝途径和建议JAK2基因有助于肺的持续的炎症反应(铍图6 b)。抑制木菠萝基因已被建议作为治疗自身免疫性炎症、木菠萝抑制剂最早成功代理达到临床应用(34,35]。JAK3 Tofacitinib (tasocitinib)抑制剂,用于银屑病,虽然TG101348,即JAK2抑制剂,对骨髓纤维化(在第一阶段试验36]。我们的研究结果表明JAK-STAT通路的潜力作为免疫调节治疗的目标CBD和其他潜在的肉芽肿性肺部疾病。
此外,我们的数据确定一个CBD血液转录签名包括TNF-α和细胞因子/趋化因子基因,在血液和认识提高监管可能导致喜神贝斯和CBD发病机理。TNF-α强有力的促炎症细胞因子,对肉芽肿的形成和维护至关重要在结核病和结节病37- - - - - -40]。米唉et al。(41)确定CBD患者增加TNF-α水平较差的临床相关疾病严重程度的标志。TNF-α* 02等位基因启动子区域是一个功能多态性与CBD严重程度(42]。我们的研究表明,CBD血液转录签名是一个强大的工具:1)洞察不同通路参与CBD的发病机制,可以进一步研究在分子水平上;2)定义潜在的治疗目标基础上额外的研究;和3)探讨差异表达基因等CCL3,CCL3L1和SOCS1这可能是重要的BeS进展。
的表达谱beryllium-stimulated BeS比CBD(类似于控制图2),这是符合我们的先前的研究结果(43]。而喜神贝斯个人演示基于BeLPT铍免疫反应,他们没有表现出显著的Th1血液中免疫反应或肺。此外,这可能是一个频谱与beryllium-exposed工人(正常健康对照组)发展敏化作用,其中一些人可能会进步CBD,或甚至可能在早期阶段的过渡。
在条款相关的生物过程,调节白细胞或T-cell-mediated细胞毒性。几乎所有的基因包括在术语编码MHC分子,如抗原,b - c, e, f, g,参与演示CD8抗原+在CBD细胞毒性t细胞。这些证据表明,CD8+t细胞也可能扮演一个角色在CBD的发展,尽管这没有评估日期。我们没有发现增加Th17基因,这并不奇怪。我们已经进行了t细胞immunophenotype研究CBD(数据进行审查),和特征的表达从患者的肺部和外周血Th17 CBD;我们发现在IL-17没有显著增加+CD4+t细胞检测到在CBD的记忆t细胞,这是符合我们当前的基因表达数据。
评价差异表达基因/通路由CBD共享和结节病显示顶部重叠网络参与炎症、免疫反应、脂质代谢和分子运输。的近300个差异表达基因之间共享CBD和结节病,STAT转录因子(STAT1和STAT2)作为一种重要的生物分子的中心连接集中(44]。有趣的是,在另一项研究中,结节病淋巴结肉芽肿表达了丰富的STAT1和p-STAT1,表明其在结节病的相关性(45]。当我们发现STAT1基因超表达PBMCs,我们还发现它增加肺泡巨噬细胞(数据没有显示)。此外,STAT1-regulated趋化因子,例如CXCL9,CXCL10,CXCL11和CXCL16在CBD和结节病病人显著调节(在线补充表S5a),表明STAT1的重要性这些疾病。STAT1可能也是一个潜在的慢性肉芽肿性疾病药物治疗的新目标。此外,只有在结节病但表达下调基因调节CBDRNASE3、编码的核糖核酸酶展品对病原菌抗菌活性。结节病可能引发的免疫系统对外来物质,如病毒、细菌或化学物质,调节的可能原因RNASE3。有可能发现CBD和结节病之间的差异可能是由于疾病本身的差异(我们假设的结果RNASE3上图),不同的隔间(PBMCs研究与肺组织),使用铍的刺激,和/或不同的病因学和/或研究设计。
除了与结节病PBMCs CBD PBMCs相比,我们比较了PBMC与肺癌基因表达和转录分析发现两者之间的差异表达基因。大部分与此同时调节基因与免疫反应(例如CCL8,CXCL9,CXCL11,TAP1,STAT1,CXCL9和CXCL11)。这些基因涉及肉芽肿形成在灵长类动物46]。有趣的是,这两个之间有19个不同的基因疾病(在线补充表S5g-f)。在结节病,但表达下调基因调节CBD是细胞大分子代谢过程(例如FYB,PSIP1,SH2D1A,CCNG2,IFNGR1和LPAR6)。唯一在结节病基因表达下调,但调节CBDRHBDD2在信号通路与细胞增殖相关的功能/生存在乳腺癌[47]。这些差异可能与这两种不同的隔间测试,一些基因是已知调节肺部或其他血液和减少(17]。这些差异可能是由于疾病严重程度的差异,尽管平均肺功能是类似的团体之间,或不同的治疗(K另外一些et al。(17包括个人治疗)。或者,这些发现可能表明,两种疾病展示一些不同方面的免疫反应和/或扩散,进一步对其进行评估。此外,我们用铍刺激血液实验揭示基因表达水平高于一些结节病的血液研究日期。此外,我们只获得transbronchial组织对CBD的诊断,这是不足以评估基因表达。我们计划研究获得BAL细胞基因表达超出了本研究的范围。
总之,我们的研究表明,喜神贝斯和CBD PBMCs展示差异表达概要文件相关的免疫功能和发病机理。此外,CBD股票与结节病致病基因/通路。本研究填补了一个关键的差距在我们目前的知识,允许新的监管基因的识别和网络,支配CBD和/或延续喜神贝斯。这个重要的转化信息可能会影响其他肉芽肿和免疫介导的疾病。最重要的是,本研究为进一步研究探索提供了基础生物标记和基因的功能和网络调查发现,最终提高我们理解CBD和其他类似疾病的发病机理。
脚注
编辑评论欧元和J2016;47:1625 - 1628。
可以从本文的补充材料www.qdcxjkg.com
支持声明:由NIH P01 ES11810-06A1, K12 HL090147-01, K01 ES020857-01和1 UL1 TR001082 NCRR /国家卫生研究院。为这篇文章一直存放在资助信息FundRef。
利益冲突:披露可以找到与这篇文章www.qdcxjkg.com
- 收到了2015年9月29日。
- 接受2016年2月7日。
- 版权©2016人队











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}