





文摘
波多黎各人在美国哮喘不成比例的影响。在这项研究中,我们的目标是识别带来对哮喘的易感性的基因变异在波多黎各人。
我们进行了一项荟萃分析的全基因组关联研究(GWAS)哮喘的波多黎各人,包括参与者:哮喘的基因在拉丁裔美国人(晚会)i ii, Hartford-Puerto Rico研究和拉美裔社区卫生研究。此外,我们研究了易感性位点是否确定在以前的荟萃分析GWAS与哮喘有关的波多黎各人。
唯一的轨迹来实现全基因组意义是17号染色体温度系数,根据我们的最高单核苷酸多态性(SNP), rs907092(或0.71,p = 1.2×10−12)IKZF3。类似于non-Puerto结果身旁,snp基因的连锁不平衡块一样IKZF3(例如ZPBP2,ORMDL3和GSDMB)在波多黎各人与哮喘相关显著。关于一个荟萃分析的结果在欧洲,我们发现为rs2305480复制GSDMB,但不是在任何其他基因snp。另一方面,我们复制的结果从一个荟萃分析的北美人口的snpIL1RL1,TSLP和GSDMB但不是IL33。
我们的研究表明,17号染色体上的常见变异温度系数在波多黎各人哮喘有最大的影响。
文摘
17号染色体上常见的等位变异温度系数对哮喘有最大的影响在波多黎各人,高危人群http://ow.ly/OtZq3084XV1
介绍
哮喘是一种与大量的遗传疾病(1,2]。全基因组关联研究(GWAS)已经确定了几个哮喘易感性位点,其中包括17号染色体区域(对方篮里3,4]。这个轨迹,其中包含的基因IKZF3,ZPBP2,GSDMB和ORMDL3,一直复制跨不同民族(3- - - - - -13]。
哮喘的负担不同种族或民族。在美国,目前哮喘是更常见的在波多黎各人(16.1%)比在非西班牙裔黑人(11.2%),非西班牙裔白人(7.7%)和墨西哥(5.4%)(14]。此外,波多黎各人从哮喘发病率比其他种族或族群的成员(15),因此在这个族群研究哮喘相关公共卫生和了解疾病的发病机理。虽然前两次GWAS哮喘包括波多黎各人参与哮喘的基因在拉丁裔美国人研究(联欢晚会我)3)和Genes-Environments和外加剂在拉丁裔美国人研究(联欢晚会II) (13),没有单独的分析提出了波多黎各人。
我们进行了一项荟萃分析GWAS的哮喘,只包括波多黎各参与者:联欢晚会,晚会II, Hartford-Puerto Rico研究(Hartford-PR) [16)和西班牙裔社区卫生研究/拉丁美洲人(HCHS /溶胶)(17,18]。此外,我们检查是否易感位点中确定两个荟萃分析GWAS的哮喘(由加布里埃尔(一个多学科研究确定哮喘的遗传和环境原因在欧洲共同体)和夏娃(识别Asthma-susceptibility基因研究在多元民族的人口)联盟(3,4在波多黎各人])与哮喘有关。
方法
研究人群
Hartford-PR
美国孩子们招募了哈特福德(CT) (n = 449)和圣胡安(波多黎各)(n = 678),其他地方报道(16]。在这两个研究地点,主要招聘工具是筛查问卷给6至14岁儿童的父母。所有参与者必须有四个波多黎各的祖父母。哮喘被定义为被诊断出的哮喘和至少一个喘息的发作。对照组在之前被诊断出的哮喘和喘息。全基因组基因分型进行了使用HumanOmni2.5 BeadChip平台(Illumina公司Inc .)、圣地亚哥、钙、美国),如前所述[16]。归责的non-genotyped单核苷酸多态性(snp)与IMPUTE2 [19),使用数据从第一阶段(2010年11月发布)1000年的人类基因组计划的参考面板。质量控制措施后,948名儿童(523哮喘)和∼700万基因分型和估算snp是包含在GWAS。使用逻辑回归分析进行了下一个加性遗传模型,调整了年龄、性别、学习网站和主成分计算使用smartPCA [20.]。写对参与孩子的父母同意了,从他书面同意也获得。这项研究是大学的机构审查委员会批准的波多黎各波多黎各的圣胡安,布莱根妇女医院(美国波士顿)和匹兹堡大学(美国宾夕法尼亚州匹兹堡)。
庆祝我
这项研究由受试者没有哮喘,招募主要来自波多黎各,但也从纽约(美国纽约)21,22]。受试者包括在这项研究如果他们8岁至40年,认同四个祖父母波多黎各。病例被诊断出的哮喘和经历过前两年的两个或两个以上的症状(气喘、咳嗽和/或气短)。对照组没有哮喘或过敏的症状,没有physician-diagnosis哮喘和无其他慢性呼吸系统疾病史或过敏(湿疹、荨麻疹或花粉热)。所有受试者基因分型Affymetrix 6.0 GeneChip(美国Affymetrix,圣克拉拉,CA),执行和质量控制所述其他地方(23]。基因分型数据是分阶段与形状(24使用IMPUTE2[执行]和归责19),使用所有的数量由1000人基因工程第一阶段v3 (25)作为参考。协会进行了测试使用逻辑回归的加性遗传模型下,调整了年龄、性别、和非洲和美洲土著祖先估计(计算使用掺合料(26])。共有437名无关的主题与哮喘(251)和完整的数据协变量对所有使用在当前分析。写对参与孩子的父母同意了,从他书面同意也获得。机构审查委员会批准的这项研究是加州大学旧金山(加州大学旧金山分校;美国旧金山,CA)和在每个参与中心。
联欢晚会二世
这是一个拉丁裔儿童哮喘病例对照研究(27]。例和对照组使用结合社区招募和诊断方法从中心在美国(芝加哥(IL),布朗克斯(纽约),休斯顿(TX),旧金山海湾地区(CA)和波多黎各)。受试者合格如果他们8到21岁,有< 10久的历史和不吸烟者,吸烟和拉丁裔种族自我报告有四个祖父母。哮喘的定义根据医生的诊断和报告症状和药物的使用在过去两年。对照组没有哮喘或过敏史,没有喘息或呼吸短促一生中。当前的分析集中在1786名参与者与哮喘(892)与自我报告的波多黎各的种族。所有受试者基因分型Axiom LAT1阵列(世界数组4;Affymetrix)和执行质量控制所述其他地方(28]。归责和协会测试了如上所述的联欢晚会。这项研究是加州大学旧金山分校的机构审查委员会批准和在每个参与中心。所有科目和他们的父母提供书面知情同意和书面知情同意。
HCHS /溶胶
的自我认同这是一个以社区为基础的群组研究18 - 74岁的拉丁美洲裔人,他们招募了在四个中心(圣地亚哥芝加哥、迈阿密、布朗克斯)。参与者招募使用两阶段抽样方案,在普查块采样单元,然后从这些单位(家庭和个人17,18]。HCHS /溶胶研究机构审查委员会批准,参与机构,并从所有参与者得到书面知情同意。
HCHS /溶胶包括那些自认为墨西哥,中美洲,南美洲、波多黎各、多米尼加和古巴。然而,遗传分析群体建立了基于这些认同拉丁美洲裔背景和遗传相似性。这些遗传分析群体很大程度上认同群体的重叠,但使用协会的遗传分析群体测试和分层分析的优势,如前所述[29日]。个人从波多黎各遗传分析组中使用当前的分析。例(n = 478)自我报告当前被诊断出的哮喘。对照组(n = 1388)从未被诊断出患有哮喘。人的基因在HCHS Illumina公司/溶胶定制15 041 502 B3数组,并估算到1000年人类基因组阶段我数据。基因分型结果的详细信息,归责和质量控制提供了其他地方(29日]。
关联分析是使用GMMAT执行(广义线性混合模型协会测试)30.),混合模式逻辑回归,占相关性由于亲缘,共享家庭和滑车组。分析调整五遗传主成分,研究中心,抽样权重(防止潜在的选择性偏差造成的抽样方案),年龄、性别、吸烟状况和久的吸烟。
在我们的主元分析,包括三个群的孩子(Hartford-PR、联欢晚会我和联欢晚会II)和一群成年人最大样本大小(HCHS /溶胶)。因为儿童哮喘可能有遗传因素,不同于那些成人哮喘,我们重复的荟萃分析排除HCHS /溶胶。
统计方法
金属(快速而有效的荟萃分析全基因组协会扫描)(31日软件是用于执行四个GWAS的荟萃分析哮喘。金属将在独立研究假定值作为输入,样本大小和作用方向考虑。首先,对于每个SNP,编码和替代确定等位基因和z分数计算基于假定值和效果在每一个研究的方向。具体来说,大积极的z分数表明小假定值的编码基因风险等位基因,和大负z分数表明小假定值编码的等位基因在哪里防护。正式的z分数是: 在哪里
在哪里 是研究的z分数吗我,
是研究的z分数吗我, 是研究的假定值吗我,
是研究的假定值吗我, 研究方向的效果吗我,
研究方向的效果吗我, 标准正态分布的百分比。然后,整个z分数和假定值计算的加权和单独的z得分:
标准正态分布的百分比。然后,整个z分数和假定值计算的加权和单独的z得分:
 在哪里Z是整体的z分数,P是整体的假定值,
在哪里Z是整体的z分数,P是整体的假定值, 是研究的重量吗我:
是研究的重量吗我: 在哪里
在哪里 是轻微的等位基因频率的研究我,
是轻微的等位基因频率的研究我, 研究病例数吗我和
研究病例数吗我和 研究控制的数量吗我。这个权重旨在密切近似的结果,将会获得结合subject-level数据研究,在一个调整的分析研究。总结优势比平均计算的一些具体log-odds比率,权重反映了标准误差的一些具体优势比。
研究控制的数量吗我。这个权重旨在密切近似的结果,将会获得结合subject-level数据研究,在一个调整的分析研究。总结优势比平均计算的一些具体log-odds比率,权重反映了标准误差的一些具体优势比。
我们使用了varLD [32,33)软件评估是否有在统计上有显著差异的地区我们的研究人群之间的连锁不平衡结构。VarLD使用蒙特卡罗方法计算假定值,它是基于000排列。两个种群的比较,p < 0.05表明连锁不平衡结构的显著差异。
结果
参与者在一年的四个特点的研究,包括荟萃分析所示表1。研究结果是基于7 485 508估算和计算基因单核苷酸多态性与哮喘和2893名对照组2144例。
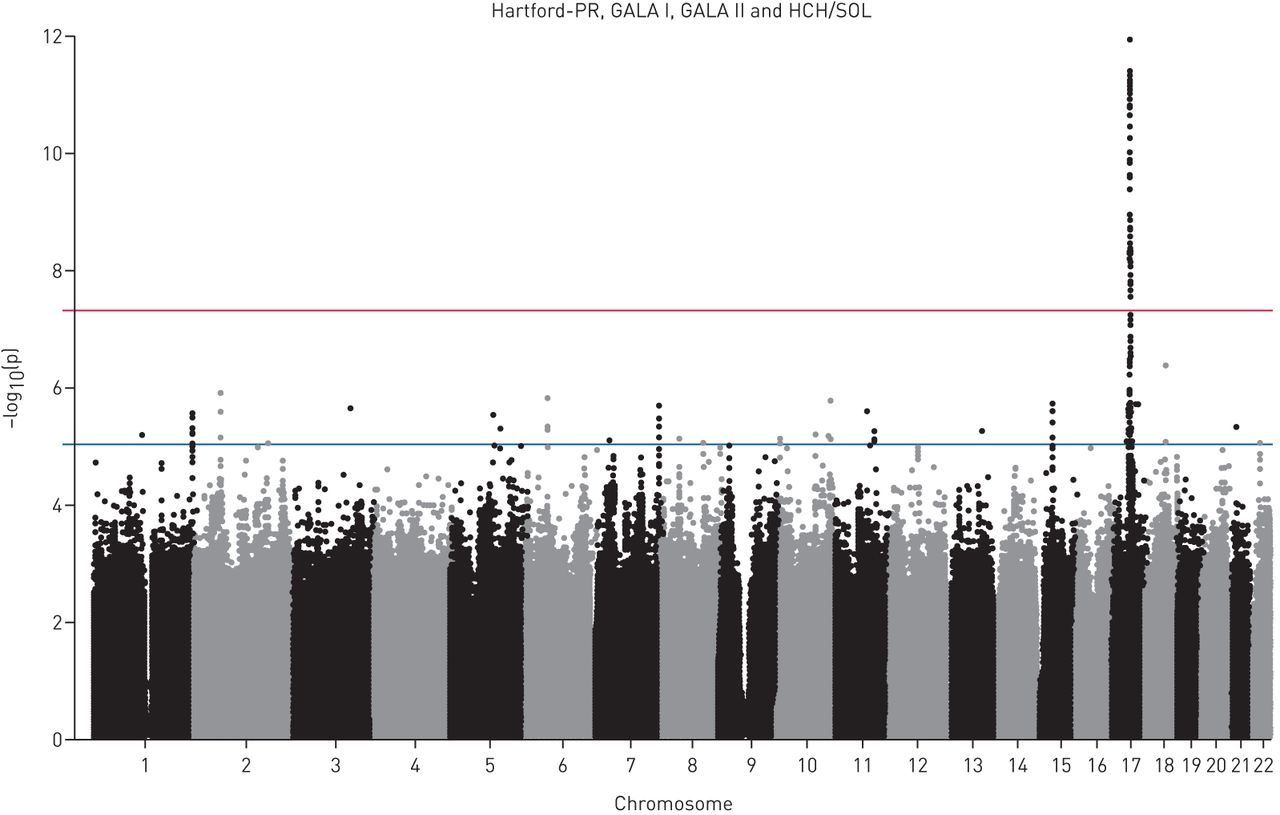
荟萃分析QQ情节(S1和S3)数据表明,无论是从每个四个组件的研究结果和合并后的结果的测试数据。此外,这些情节透露大量的小假定值研究相结合。我们将看到单核苷酸多态性与假定值< 5×10−8,标准的意义对GWAS阈值。在合并后的研究中,我们发现,17号染色体上的89个snp温度系数是重要的在这个级别(图1和表S1)。这些89个snp, rs907092IKZF3显示最重要的协会与哮喘(p = 1.2×10−12,图2和3)。这个SNP也与哮喘有关p < 0.023在所有这三个研究的儿童和在HCHS p = 0.092 /溶胶,成年人的研究(图2和图S6)。
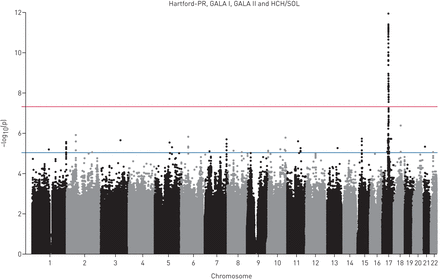
曼哈顿的情节显示Hartford-PR的总结分析结果,联欢晚会,晚会二世和HCHS /溶胶。显示每个SNP的染色体位置沿x轴和协会的负对数假定值显示在y轴。蓝线代表暗示意义(p < 1×10行−5)。红色的线条代表的全基因组意义(p < 5×10行−8)。
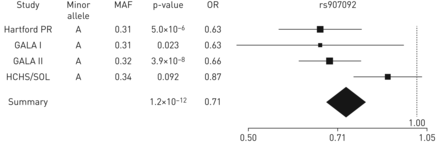
森林土地的优势比和95%置信区间与哮喘。森林rs907092情节,荟萃分析中最重要的SNP。
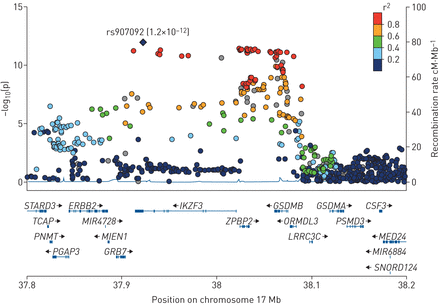
荟萃分析的结果(Hartford-PR联欢晚会,晚会II和HCHS /溶胶)在17号染色体区域。基因和转录的方向的相对位置较低部分的图所示,和染色体的位置显示在x轴。浅蓝色的线显示了该地区的重组率(右y轴),离开了y轴显示的意义关联。紫色的钻石显示了rs907092的假定值最重要的SNP的荟萃分析(Hartford-PR,联欢晚会我晚会II和HCHS /溶胶)。圆圈显示所有其他snp的假定值和颜色编码根据LD的水平与rs907092 1000基因组计划混在美国人口(AMR)。
鉴于我们17号染色体结果温度系数和哮喘,尤其是在研究包括儿童,我们检查了这个地区的连锁diseuilibrium模式在Hartford-PR波多黎各人,联欢晚会我和联欢晚会。S8-S10数据显示,受试者在这些研究中也有类似的连锁不平衡模式(varLD Hartford-PR假定值= 0.08与晚会我;Hartford-PR假定值= 0.46与联欢晚会II(表S2)),几乎所有重要的snp聚集在一个连锁不平衡本跨越∼200 kb(参见图S8-S10)。的IKZF3-ZPBP2-GSDMB-ORMDL3轨迹包含在这一本。然后我们17号染色体的连锁不平衡模式相比温度系数在波多黎各人与其他民族(图S11)。波多黎各人有连锁不平衡模式,不同于在墨西哥人或欧洲人(varLD假定值Hartford-PR≤0.0007与墨西哥人(我在晚会或联欢晚会II)或欧洲人(表S2) 1000克)。墨西哥基因型分析,提取从晚会我和二联欢晚会,和欧洲从1000人基因工程中提取基因型。
接下来,我们检查了单核苷酸多态性是否之前在一个荟萃分析与哮喘有关的GWAS的欧洲人(4]或种族多样化的北美人口的荟萃分析(3波多黎各人)也与哮喘相关(表2)。荟萃分析的欧洲人,我们能够复制协会的SNPGSDMB在p < 0.005 (Bonferroni调整为10个测试)。荟萃分析的多元民族的人口,我们能够复制与单核苷酸多态性IL1RL1,TSLP和GSDMB在p < 0.01 (Bonferroni调整为5测试)。这两个单核苷酸多态性(rs2305480和rs11078927)GSDMB确定两个引用研究一直在Hartford-PR假定值小,联欢晚会,晚会,以及在荟萃分析。我们的SNP (rs907092)IKZF3在高LD (r2= 0.84 - -0.85)与rs2305480 (SNP在报道GSDMB从加布里埃尔(一个多学科研究来识别基因和环境引起的哮喘欧洲共同体))和高LD (r2= 0.84 - -0.85)与rs11078927 (SNP在报道GSDMB从夏娃(识别Asthma-susceptibility基因研究在多元民族的人口))。r2rs2305480与rs11078927大于0.99。我们没有复制协会与单核苷酸多态性IL33报道,由以前的荟萃分析。因为这可能是由于不同的LD模式或附近IL33波多黎各和族群之间的包含在之前的荟萃分析(如。欧洲人和墨西哥人;图S12),我们也显示IL33表S3中snp的荟萃分析。在这个分析中,没有IL33SNP与哮喘相关显著(p > 0.00035在所有情况下,Bonferroni调整为142测试)。
因为儿童哮喘可能有遗传因素,不同于那些成人哮喘,我们重复GWAS的荟萃分析哮喘没有HCHS /溶胶,获得相似的结果(数字S4, S5和S7)的荟萃分析包括HCHS /溶胶:只在17号染色体snp温度系数达到了显著性水平p < 5×10−8。此外,我们能够复制相同的snp从GWAS的前两个荟萃分析哮喘的荟萃分析包括HCHS /溶胶(表S4)。此外,单核苷酸多态性IL18R1和GSDMA复制在p < 0.005 (Bonferroni调整为10个测试)。
为了检查是否有独立的单核苷酸多态性与哮喘有关,我们17号染色体进行条件分析温度系数通过调整顶部SNP在我们分析(rs907092)。由于成年人没有表现出强烈的遗传协会在这个地区,我们进行了条件分析的荟萃分析三个群的孩子。结果表明,没有17号染色体snp在到达对方篮里Bonferroni——纠正显著性水平α= 4.1×10−5(1215重叠snp与小等位基因频率(加)> 0.05在三组)和没有snpIKZF3-ZPBP2-GSDMB-ORMDL3轨迹到达Bonferroni-corrected显著性水平α= 2.5×10−4(197重叠snp与加跨三个军团> 0.05)。因此,除了单核苷酸多态性与rs907092连锁不平衡,似乎没有其他独立信号的染色体17温度系数。
讨论
我们报告的发现从第一GWAS哮喘进行单独在波多黎各人。虽然两组包含在目前的荟萃分析(联欢晚会我和II)是拉丁美洲裔人群的一部分包含在先前的研究,这些研究没有分别从波多黎各人,而是结合数据分析数据来自波多黎各和墨西哥人。这种方法可以减少统计力量,因为波多黎各人,墨西哥人明显对不同种族血统(即。波多黎各人,平均而言,更大比例的非洲血统,但较低的比例比墨西哥印第安人血统),哮喘负担(如墨西哥人有哮喘的负担远低于波多黎各人)和等位基因频率(15]。
荟萃分析的四个GWAS,唯一的轨迹实现全基因组重要协会17号染色体与哮喘的温度系数,根据我们的SNP, rs907092 (p = 1.2×10−12),位于IKZF3。类似于non-Puerto之前发现身旁,单核苷酸多态性的基因位于同一连锁不平衡块IKZF317号染色体上温度系数(例如ZPBP2,ORMDL3和GSDMB)也显著与哮喘相关的荟萃分析。前一个候选基因研究在399年波多黎各和301年墨西哥人在联欢晚会我,以及261非洲裔美国人,发现重大的两个单核苷酸多态性之间的关联ORMDL3(rs4378650和rs12603332)和哮喘在墨西哥人和非裔美国人,但不是在波多黎各人对单核苷酸多态性(p = 0.08) [6]。在我们的分析中,这两个单核苷酸多态性(p = 4.0×10−6rs4378650和p = 5.5×10−6rs12603332)被复制和哮喘的关系。
与之前研究结果一致表明强影响17号染色体snp的温度系数比在成人哮喘儿童哮喘在欧洲(4),我们发现全基因组重要结果这轨迹分析局限于三个群的孩子,但不是在成年人哮喘的GWAS HCHS /溶胶。
我们也试图复制前发现潜在asthma-susceptibility snp在欧洲进行的荟萃分析(GABRIEL财团)[4)和种族多元化的北美人口(夏娃财团)[3]。从加布里埃尔对于结果,我们复制结果的SNPGSDMB(17号染色体上温度系数),但不是任何其他基因的snp。另一方面,我们报告的第一个复制的结果多民族夏娃财团的snpIL1RL1,TSLP和GSDMB在一个队列由专门的波多黎各人。我们缺乏加布里埃尔在波多黎各人模仿复制的结果负面结果前夕财团,和可能是由于民族差异风险变异哮喘。事实上,夜17号染色体只复制发现对方篮里IL33从加布里埃尔(尽管不同的单核苷酸多态性)。
我们没有复制的结果PYHIN1或IL33从夏娃。PYHIN1只是与哮喘相关的非裔美国人,非洲-加勒比裔,和血统的差异也许可以解释我们的负面结果。单核苷酸多态性与加≥0.37(加IL33SNP rs2381416在我们的研究人群),我们统计≥70%功率检测的优势比≥1.10α≤0.01我们的荟萃分析34]。在同样的假设,统计能力检测的优势比≥1.20α≤0.01是我们荟萃分析> 99%。的142个snpIL33是检测与哮喘在最近的研究中,只有6个相关,p < 0.01,没有Bonferroni调整后仍与哮喘相关显著。
表达式的17个基因,对方篮里ORMDL3,是由asthma-associated单核苷酸多态性(35),复制在我们之前的表达式(eQTL)研究数量性状(rs8067378, p = 2.6×10−10)使用Hartford-PR数据36]。最近,ORMDL3涉及多种细胞过程,可能与哮喘相关(35]。除了对哮喘易感性的影响,研究从Tavendaleet al。(11)还显示,ORMDL3与哮喘急性加重,ORMDL3SNP-mediated表达式是影响鼻病毒感染,常见的引发的急性加重(37]。苏格兰民族党在IKZF3-ZPBP2-GSDMB-ORMDL3地区与哮喘有关的夏娃荟萃分析rs11078927在GSDMB复制在我们的研究中(p = 8.1×10−12(表2))。前夕,这个SNP和哮喘的报道优势比分别为0.80 0.78在欧洲美国和拉丁美洲人(非重要非裔美国人)。在我们的研究中,我们估计,所有科目优势比0.72和0.66的儿童,暗示的强大效果IKZF3-ZPBP2-GSDMB-ORMDL3轨迹在儿童哮喘波多黎各人比其他种族的人。因此,主要的影响等位基因SNP rs11078927 (即。或1.51(1/0.66))可能导致儿童哮喘的病因学在波多黎各人。
的ZPBP2-GSDMB-ORMDL3轨迹检测中由于等位基因的差异表达差异lymphoblastoid细胞系(拼箱)和CD4+细胞(38,39]。早期研究基因表达的40- - - - - -42)透露,asthma-associated snp规范的表达ORMDL3和GSDMB,和这两个基因可能coregulated [38]。发现之前的一项研究表明,一个asthma-associated 17对方篮里监管单体型影响转录活性ZPBP2, GSDMB和ORMDL3,非编码在ZPBP2变体,rs12936231,不同影响绝缘子的绑定蛋白CTCF allele-specific地(38];随后eQTL研究表明,SNP rs12936231增加ORMDL3 GSDMB表达式在初级淋巴细胞,全血和肺组织43,44]。然而,SNP rs12936231(一个小等位基因频率= 0.44 - -0.46在我们组)与哮喘在荟萃分析与相关(p = 2.0×10−6),只是在温和的连锁不平衡(r2> 0.51)与我们的SNP (rs907092)IKZF3。这表明,17号染色体上的其他功能多态性可能影响对方篮里哮喘风险在波多黎各人,可能通过改变表达ORMDL3(如图所示,导致转基因小鼠实验性哮喘模型)(45),GSDMB或ZPBP2。另一个SNP位于启动子区域ZPBP2(rs4795397, p = 3.7×10−12在我们的荟萃分析(表S1))是一个假定的功能多态性显示asthma-associated allele-specific核小体占用(39]。然而,苏格兰民族党的强烈影响ZPBP2推广活动是蒙面的DNA甲基化基因外显子1。相比之下,ORMDL3启动子是完全unmethylated。的ZPBP2和ORMDL3显示等位基因的差异表达(38,39]。它已被证明IKZF3-ZPBP2-GSDMB-ORMDL3单体型不同行为的监管记录之间的欧洲人和非洲人(38),与一个更强大的协会在欧洲比在非洲人。在一项由Verlaanet al。(38),SNP rs8067378是最重要的独联体-eQTL在这个基因研究的四个地区的欧洲人(p = 1.1×10−18)和非洲(p = 4.3×10−9)。在波多黎各人使用我们的Hartford-PR eQTL研究数据集(36),同样的SNP也是最重要的独联体-eQTL (p = 2.6×10−10)在基因研究的四个地区。尽管这两项研究[36,38)可能不是由于不同的统计模型和不同的组织(即。,拼箱用于Verlaanet al。(38)研究和globin-cleaned全血中使用C母鸡et al。(36]研究),我们推测的意义相关联的单核苷酸多态性与表达之间的波多黎各人是欧洲人和非洲人,因为这是两个波多黎各人的祖先种群。我们进一步eQTL执行分析的SNP (rs907092)和表达IKZF3-ZPBP2-GSDMB-ORMDL3使用遗传核糖核酸微阵列数据轨迹从Hartford-PR研究(n = 121)。从这二次分析结果,进行了使用线性回归和调整了年龄、性别、哮喘状态,学习网站和前两个主成分,表明我们SNP (rs907092)可能的规范表达IKZF3-ZPBP2-GSDMB-ORMDL3轨迹(p = 0.07IKZF3,p = 0.02ZPBP2,p = 0.009GSDMB和p = 0.0004ORMDL3)。
我们认识到几个研究的局限性。首先,我们有统计能力不足检测弱常见snp遗传效应(如。优势比0.9和1.10之间IL33)或罕见的敏感性变异。第二,学科在我们的研究在不同的网站在美国大陆和波多黎各,因此环境差异可能会混淆我们的结果尽管调整研究网站的四个组件的研究。然而,大多数小等位基因频率并在研究群体效应估计是相似的。第三,错误分类的慢性阻塞性肺病尽可能哮喘是成年人HCHS /溶胶队列,鉴于980年(52.5%)参与者前任或现任吸烟者(意味着久的18.6)。这样的误诊可能占显著关联报道HCHS /溶胶越少,即使调整吸烟。总之,我们的研究表明,常见的等位基因变异在17号染色体位点对方篮里有最大的影响,因此也特别重要,在波多黎各人哮喘的发病机制。我们证实敏感性变异两个之前报道基因(IL1RL1和TLSP)。未来的研究应该致力于描述功能变异导致哮喘在波多黎各人,通过主基因效应或与相关风险敞口(交互如。二手烟和空气污染)。
补充材料
披露的信息
确认
我们感谢Hartford-PR研究的参与者和员工,联欢晚会我和联欢晚会II, HCHS /溶胶为这项工作研究的贡献。
作者的贡献:概念和研究设计:R.G.巴尔,今后如J.C.青瓷;数据采集、分析和解释:问:燕,m . Pino-Yanes j . Brehm e .《j .林号哦,e . Acosta-Perez林祖嘉劳丽,M.M.克劳蒂尔在,文学士学位雷比,上午Stilp, t .开发d . Hu s洪博培C.S. Eng,议员Conomos, d . Rastogi k .大米、g . Canino和w·陈;和起草手稿的知识内容:问:燕,R.G.巴尔,如伯查德和J.C.青瓷。所有作者批准之前提交最终版本的手稿。
脚注
可以从本文的补充材料www.qdcxjkg.com
支持声明:这项工作是HL079966赠款支持和HL117191来自美国国立卫生研究院(NIH)和亨氏禀赋。问:燕的贡献被授予支持UPMC的匹兹堡儿童医院。e .《的贡献被授予HL125666来自美国国立卫生研究院支持。d . Rastogi NIH的HL118733支持的贡献。联欢晚会II今后研究是由美国国家卫生研究院的基金如:HL088133, HL004464, HL117004, ES015794, ES24844, TRDRP 24 rt 0025年MD006902, GM007546。今后如也投票支持的阿莫斯医学院开发奖、美国哮喘基金会和桑德勒基金会。m . Pino-Yanes从学院祝您健康支持格兰特AC15/00015卡洛斯三世在ERACoSysMed 1日联合跨国电话从欧盟(SysPharmPedia 99), 2020年在地平线下。HCHS /溶胶是由国立卫生研究院合同N01-HC65233, N01-HC65234, N01-HC65235, N01-HC65236, N01-HC65237, HHSN268201300005C AM03。基线检查HCHS /溶胶进行协作研究支持的合同从国家心脏,肺和血液研究所(NHLBI)到北卡罗莱纳大学(N01-HC65233),迈阿密大学(N01-HC65234),爱因斯坦医学院(N01-HC65235),西北大学(N01-HC65236)和圣地亚哥州立大学(N01-HC65237)。下面的研究所/中心/办公室造成的第一阶段HCHS /溶胶NHLBI通过转移基金:国家人群健康状况及风险研究所,国家耳聋和其他沟通障碍研究所,国家牙科和颅面研究所(NIDCR),国家糖尿病、消化和肾脏疾病研究所,国家神经疾病和中风研究所NIH Institution-Office膳食补充剂。 The Genetic Analysis Center at the University of Washington was supported by NHLBI and NIDCR contracts (HHSN268201300005C AM03 and MOD03). Funding information for this article has been deposited with theCrossref资助者注册表。
利益冲突:披露可以找到与这篇文章www.qdcxjkg.com
- 收到了2016年7月27日。
- 接受2016年12月18日。
- 版权©2017人队











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}