





文摘
支气管旁血管生成可能发生在囊性纤维化和血管内皮生长因子(VEGF)——调节血管生成在航空公司。
支气管旁多血管和VEGF-A表达检测采用免疫细胞化学和肺的形态学分析部分获得10囊性纤维化患者移植与10控制不吸烟者,在两个Cftr-deficient小鼠与野生型菌株的同胞。气道上皮细胞NCI-H292 noncystic纤维化的主要文化人类呼吸道上皮细胞治疗与囊性纤维化跨膜电导调节(雌性生殖道)抑制剂(CFTR-inh172年ppq - 102)或转染雌性生殖道小干扰RNA (si)有或没有选择性表皮生长因子受体酪氨酸激酶抑制剂。VEGF-A和磷酸化表皮生长因子受体的浓度由ELISA测定。
支气管旁多血管增加在囊性纤维化患者中,但不是在雌性生殖道有缺陷的老鼠。VEGF-A疣状是局部的气道上皮和囊性纤维化患者和增加雌性生殖道有缺陷的老鼠。在人工气道上皮细胞,治疗与雌性生殖道抑制剂或转染小干扰rna诱导雌性生殖道VEGF-A双重的增加产量。雌性生殖道抑制剂引起的表皮生长因子受体磷酸化VEGF-A合成所需。
囊性纤维化航空在移植显示增加支气管旁多血管和上皮VEGF-A表达式。VEGF-A雌性生殖道功能障碍引起的上皮细胞合成,这可能导致血管重塑。
文摘
VEGF-A雌性生殖道功能障碍引起上皮细胞合成,这可能导致血管重塑http://ow.ly/p7ebY
介绍
囊性纤维化(CF)是一种遗传性疾病引起的突变在囊性纤维化跨膜电导调节(雌性生殖道)基因1]。雌性生殖道异常蛋白质功能障碍导致离子运输在气道上皮细胞(2),逐步导致慢性肺疾病(1,3]。CF肺部疾病的特点是与粘液堵塞气道结构异常,包括支气管扩张,牙龈纤维化,气道上皮和粘膜下腺体的改造,气道平滑肌增生(4- - - - - -6]。
支气管循环能够增殖并支气管旁血管生成(新血管的增殖从现有船只)也正越来越被认为是一个重要的组件在慢性气道疾病(7,8]。异常支气管旁血管可能导致免疫反应通过微血管渗漏和招聘的炎症细胞(9,10]。增加支气管旁多血管被报道在哮喘和慢性阻塞性肺疾病(7,8]。然而,我们所知,支气管旁多血管没有检查在CF病理研究。
血管内皮生长因子(VEGF)——是一个关键的血管生成中介在航空公司8]。Baluket al。(11]表明,选择性的表达VEGF-A在小鼠气道上皮细胞引起气管血管生成。先前的研究在CF主题报道增加VEGF-A血清中浓度(12- - - - - -14),但对VEGF-A表达CF航空公司。Krennet al。(15)发现VEGF-A信使rna和蛋白质增加肺匀浆的CF受试者接受肺移植。相比之下,米注视者et al。(13)没有发现VEGF-A水平增加支气管肺泡灌洗液的成年CF科目。因此,表达式模式和细胞本地化VEGF-A CF航空公司仍有待建立。
在目前的研究中,我们检测支气管旁多血管和VEGF-A疣状肺组织部分从CF科目与控制不吸烟者的肺,两种不同的菌株雌性生殖道有缺陷的老鼠与野生型的同胞。由于气道上皮细胞出现的主要网站VEGF-A CF航空和表达式雌性生殖道不足的老鼠,我们研究了雌性生殖道功能障碍的影响在培养VEGF-A合成气道上皮细胞。
方法
人类和小鼠的组织
外围人类肺组织(包含noncartilaginous航空公司)在移植获得从10不吸烟的CF从10不吸烟的成年人和控制接受外围肺癌肺切除的。这些患者的临床特征提供了在线补充表S1。肺组织样本固定在10%中性缓冲福尔马林inflation-immersion和嵌入在石蜡。减少潜在的抽样偏差相关的CF患者不规则分布的形态异常,组织了从2到4块(取决于组织的可用性),其位置是随机抽取的。在对照组中,只有一个块被用于研究。
雌性生殖道m1UNC(雌性生殖道- / -)小鼠,建立了基因打靶(16)及其野生型的同胞(雌性生殖道+ / +)从中心分布,获得德Typage et d 'Archivage动物(法国奥尔良)。雌性生殖道tm1Eur(F508▽/▽)的老鼠17)及其野生型的同胞(F508wt / wt从b)老鼠一种礼物他(伊拉斯姆斯大学鹿特丹,荷兰)。断奶后,所有老鼠提供了一个商业渗透泻药(Movicol;英国Harefield Norgine)饮用水,防止肠道阻塞。8 - 10周大的老鼠牺牲和肺在4%福尔马林固定石蜡和嵌入。动物研究进行5 - 7每组。这项研究符合《赫尔辛基宣言》和当地所有的规则委员会对人类和动物研究。从所有患者知情同意了。
免疫组织化学染色和定量的形态学分析
免疫组织化学染色进行5-μm石蜡包埋部分,如前所述[18]。以下主要使用抗体:多克隆抗体VEGF-A (20, 1:200;圣克鲁斯生物技术有限公司、圣克鲁斯、钙、美国);多克隆抗体的内皮标记血管性血友病因子(vWF 1:50;美国DakoCytomation Carpinteria, CA)来识别血管。抗原暴露与蛋白酶从孵化链霉菌属将(0.01%;Sigma-Aldrich有限公司、吉林厄姆、英国)需要20分钟为vWF提高染色质量。生物素化的anti-rabbit抗体(1:200;向量实验室,伯林盖姆、钙、美国)是用于二次抗体,结合抗体呈现是根据标准协议avidin-biotin-peroxidase复杂方法(ABC精英设备;向量的实验室,英国彼得伯勒)。组织部分与苏木精复染色。遗漏的初级抗体和孵化与不相关的免疫球蛋白作为消极的控制。
量化的支气管旁多血管和VEGF-A上皮疣是由点计数,如前所述[4,18,19]。卷被VEGF-A积极染色细胞上皮细胞是上皮细胞的表达每个卷。支气管旁多血管被定义为体积被vWF-positive血管支气管旁空间之间的空间(定义为肺泡上皮基底膜和基地附件)每卷的支气管旁空间。所有分析至少20个随机选择的高倍率(400×)显微照片获得在遥远两到四个部分每病人或鼠标。
人类文化气道上皮细胞
NCI-H292人工气道上皮细胞(HAECs)表达CFTR功能检测(20.)和生产VEGF-A (18),镀在105细胞·毫升−1在24-well板块(BD猎鹰;美国BD生物科学,圣何塞,CA),和生长在RPMI 1640中含10%胎牛血清(FCS)、青霉素(100 U·毫升−1)、链霉素(100μg·毫升−1)和消息灵通的(25毫米)37°C湿润5%二氧化碳水套孵化器。
的主要文化HAECs得到从五个non-CF鼻息肉患者接受手术,如前所述[21]。细胞被镀在106细胞·好−1在插入(12毫米;Transwell合演,剑桥,美国MA)与12-mm-diameter IV型collagen-coated (Sigma-Aldrich)聚碳酸酯微孔膜和孵化1毫升的DMEM / F12 /抗生素含有2% Ultroser G (Gibco BRL,生活技术,Gaithersberg,医学博士,美国)。细胞在气液界面培养3周,每次当他们显示完整的分化与纤毛粘液细胞(22]。之前的研究表明,这些细胞:1)生产VEGF-A蛋白生产高峰在第一周的文化,文化和减少生产之后,直到天21 (21];和2)没有雌性生殖道的表达和功能在第一周的文化,但文化表达CFTR功能检测2周后(23]。因此,研究使用雌性生殖道抑制剂(见下文)进行21天的文化。
在雌性生殖道抑制HAECs
抑制CFTR-mediated离子运输是通过治疗和选择性雌性生殖道抑制剂(CFTR-inh气道上皮细胞172年或ppq - 102;Calbiochem,诺丁汉,英国)24,25),据报道,稳定雌性生殖道频道在一个封闭的状态。总之,在60 - 80% NCI-H292细胞融合与CFTR-inh接受治疗172年(10−5米)或ppq - 102 (10−5M)稀释在二甲亚砜(DMSO),或DMSO(σ)72 h。曝光时间和抑制剂的浓度是根据以往的文献[选择24- - - - - -26]。在初步实验中,我们发现,雌性生殖道抑制剂诱导VEGF-A合成细胞血清饥饿时(FCS)在过去的48 h(文化和细胞培养时的FCS(数据未显示);所有后续实验中执行FCS-containing媒介。因为HAECs在气液界面培养,因为雌性生殖道表达在上皮细胞的顶端,治疗应用在基底外侧水平(培养基),和在顶端水平下降150μL培养基在细胞表面(26]。文化媒体每天都在变化。72 h后孵化与雌性生殖道抑制剂或DMSO,细胞培养媒体收集和储存在-80°C。VEGF-A浓度,对应VEGF-A生产24 h,通过ELISA测定(见下文)。
击倒的雌性生殖道的表达是通过使转染NCI-H292细胞与特定的雌性生殖道小干扰RNA(σ),(si)和小干扰RNA作为负控制不属预定目标的27]。使用TransIT-siQUEST1转染进行,根据制造商的说明(WI Mirus,麦迪逊,美国)27]。由我们组报道在之前的手稿(27),雌性生殖道siRNA导致60 - 70%击倒在雌性生殖道表达式中,所研究的免疫印迹(数据没有显示)。转染24 h后,细胞被冲洗三次与新鲜培养基和孵化24小时之前收获文化媒体,是储存在-80°C。
在选定的实验中,上皮细胞被孵化24 h和选择性表皮生长因子受体(EGFR)抑制剂(AG1478 10−5M;Calbiochem)或其活性类似物(AG9 10−5M;Calbiochem) [28]。评估雌性生殖道的作用抑制表皮生长因子受体激活,NCI-H292细胞CFTR-inh对待172年或DMSO仅15分钟,1 h, 3 h, 6 h, 8 h和24 h。在每个时间点,细胞被洗两次无菌磷酸盐和细胞溶解产物通过溶解细胞在107细胞·毫升−1溶菌产物缓冲区(1% NP-40替代(圣克鲁斯生物技术、海德堡、德国),20毫米三羟甲基氨基甲烷在液pH值8.0,137毫米氯化钠,10%的甘油,EDTA 2毫米,1毫米激活钠原钒酸盐,μg·10毫升−1抑肽酶和10μg·毫升−1亮抑酶肽)。样品被允许坐在冰前15分钟存储在-80°C。
VEGF-A和磷酸化EGFR蛋白浓度的测量
VEGF-A在细胞培养上清液和磷酸化表皮生长因子受体(EGFR-P)在细胞溶解产物被夹心ELISA试剂盒测量根据制造商的指示(DuoSet IC;研发系统,明尼阿波利斯,美国)。EGFR-P结果表示为pg的EGFR-Pμg总蛋白质。蛋白质浓度在细胞溶解产物测定使用bicinchoninic酸碱度蛋白质分析工具包(美国皮尔斯,罗克福德,IL)。每个样品重复测定。
定量PCR对VEGF mRNA
从NCI-H292细胞总RNA提取使用RNeasy迷你包(试剂盒、希尔登,德国)。1μg总RNA用于逆转录定量PCR (qPCR)放大后VEGF-A和β-actin表达式使用7900 ht快速实时PCR系统(应用生物系统公司;生命科技有限公司,英国佩斯利)。qPCR引物被用作VEGF-A: 5′-AGGGCAGAATCATCACGAAGT-3′(向前),5′-AGGGTCTCGATTGGATGGCA-3′(反向),以及β-actin: 5′-GCAAACGTGCGTGACAT-3′(向前),5′-TCGCGACAGGTCTTTGC-3′(反向)。基因表达的相对量化ΔΔCt计算方法和结果是成倍增加在每个时间点与相应的控制。
统计分析
数据获得的形态学分析提出了%和使用非参数分析Mann-Whitney u测验和斯皮尔曼等级相关。interobserver系数变化的形态学尺寸< 15%。从细胞培养实验中获得的数据被当作意味着±扫描电镜至少有三个独立的实验中执行复制并使用单向重复测量的方差分析,分析的紧随其后事后Student-Neuman-Keuls测试多个比较。所有分析使用棱镜5软件(GraphPad软件公司,圣地亚哥,美国)。假定值≤0.05被认为是显示统计学意义。
结果
在人类和小鼠支气管旁多血管和VEGF-A疣状航空公司
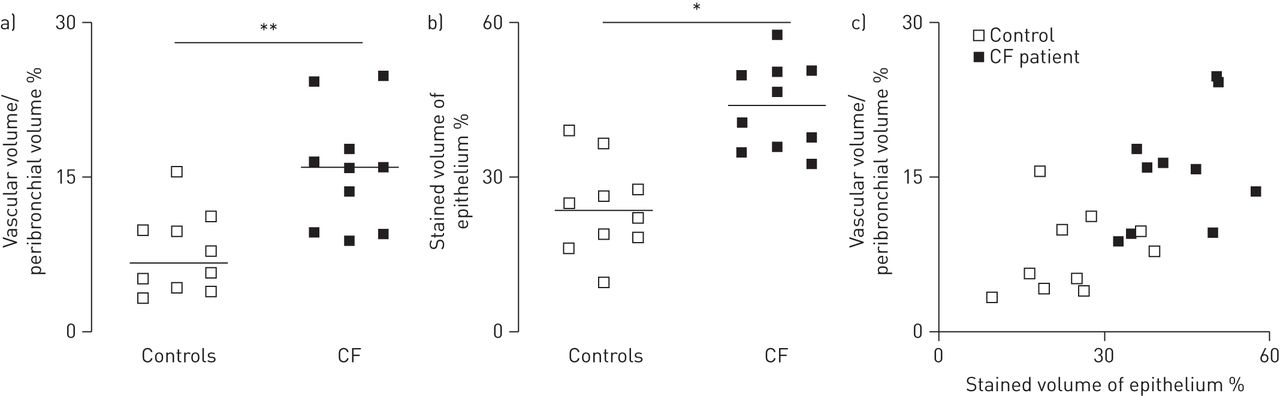
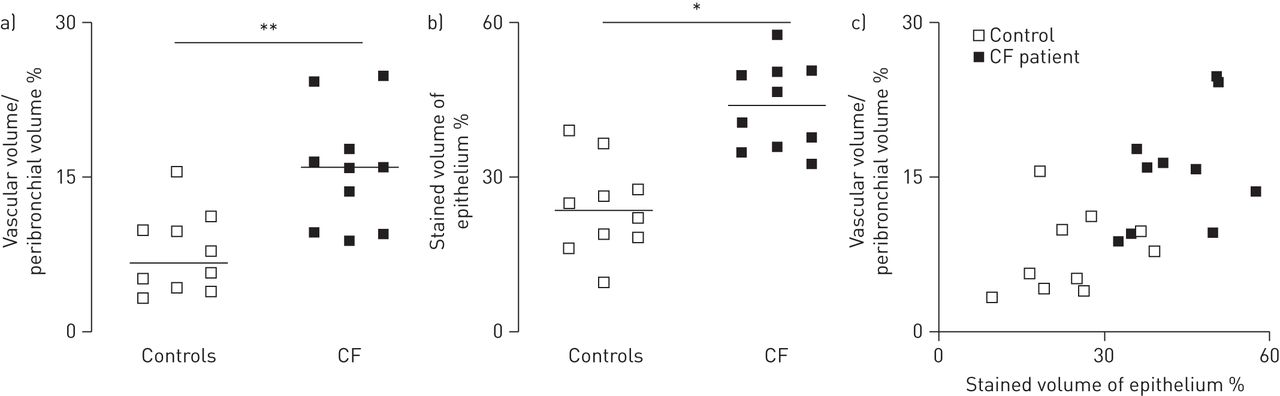
控制不吸烟者,支气管旁空间薄和疣状vWF识别稀疏支气管旁血管。CF学科,支气管旁空间显著增厚,含有大量的血管。的形态学分析显示支气管旁多血管在CF航空公司增加了两倍(p < 0.01) (图1一个)。疣状VEGF-A是积极的在肺段的控制和CF科目。VEGF-A染色是主要是局部的气道上皮细胞,存在于纤毛,但不是在杯状细胞。在CF航空,VEGF-A染色也存在于巨噬细胞,但不是在中性粒细胞(没有显示)。的形态学分析表明,VEGF-A染色明显增加气道上皮的CF科目(p < 0.05) (图1 b)。当分组控制和CF,很强的相关性之间的上皮VEGF-A-stained体积密度和支气管旁血管体积密度被发现(r = 0.65;p < 0.05) (图1 c)。然而,这种相关性不显著,当限制分析CF科目(r = 0.45;p = 0.19)。
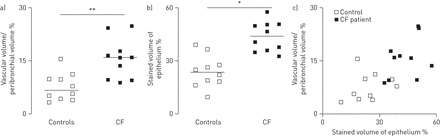
的形态学分析支气管旁多血管和血管内皮生长因子(VEGF)——免疫染色的航空管制(n = 10)和囊性纤维化(CF)的科目(n = 10)。气道部分应用了内皮标记血管性血友病因子(识别血管)或VEGF-A。的形态学分析使用点计数(见方法部分)。)显示了支气管旁血管,定义为体积被血管(血管体积)支气管旁支气管旁空间的空间/卷(%)。b)显示了VEGF-A积极上皮染色的体积百分比。c)显示了血管体积/支气管旁体积百分比和上皮染色VEGF-A积极作为的体积百分比。相关性VEGF-A上皮和支气管旁多血管都使用了非参数斯皮尔曼等级相关。这种相关性非常显著(r = 0.65;p < 0.05)在考虑控制和CF的主题,但不是重要的在考虑只有CF科目(r = 0.45;p = 0.19)。 Horizontal bars represent medians. *: p<0.05; **: p<0.01.
在雌性生殖道- / -和F508▽/▽小鼠和野生型小鼠,疣状VEGF-A也是局部的气道上皮细胞。的形态学分析显示VEGF-A-staining上皮细胞的增加雌性生殖道- / -和F508▽/▽与各自的野生型小鼠相比同窝出生的(图2)。没有区别支气管旁多血管比较时被发现雌性生殖道不足和野生型小鼠(数据没有显示)。
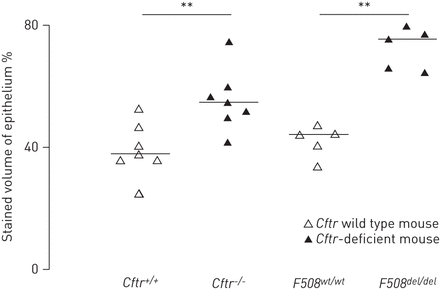
血管内皮生长因子(VEGF)的形态学分析——气道上皮的疣状囊性纤维化跨膜电导调节(雌性生殖道)缺乏和野生型老鼠。气道部分获得雌性生殖道m1UNC(雌性生殖道- / -)小鼠和野生型(雌性生殖道+ / +)同窝出生(左),雌性生殖道tm1Eur(F508▽/▽)及其野生型(F508wt / wt)同窝出生(右)。部分应用了VEGF-A和形态学分析方法中所描述的部分。单杠表示中位数。* * p < 0.01。
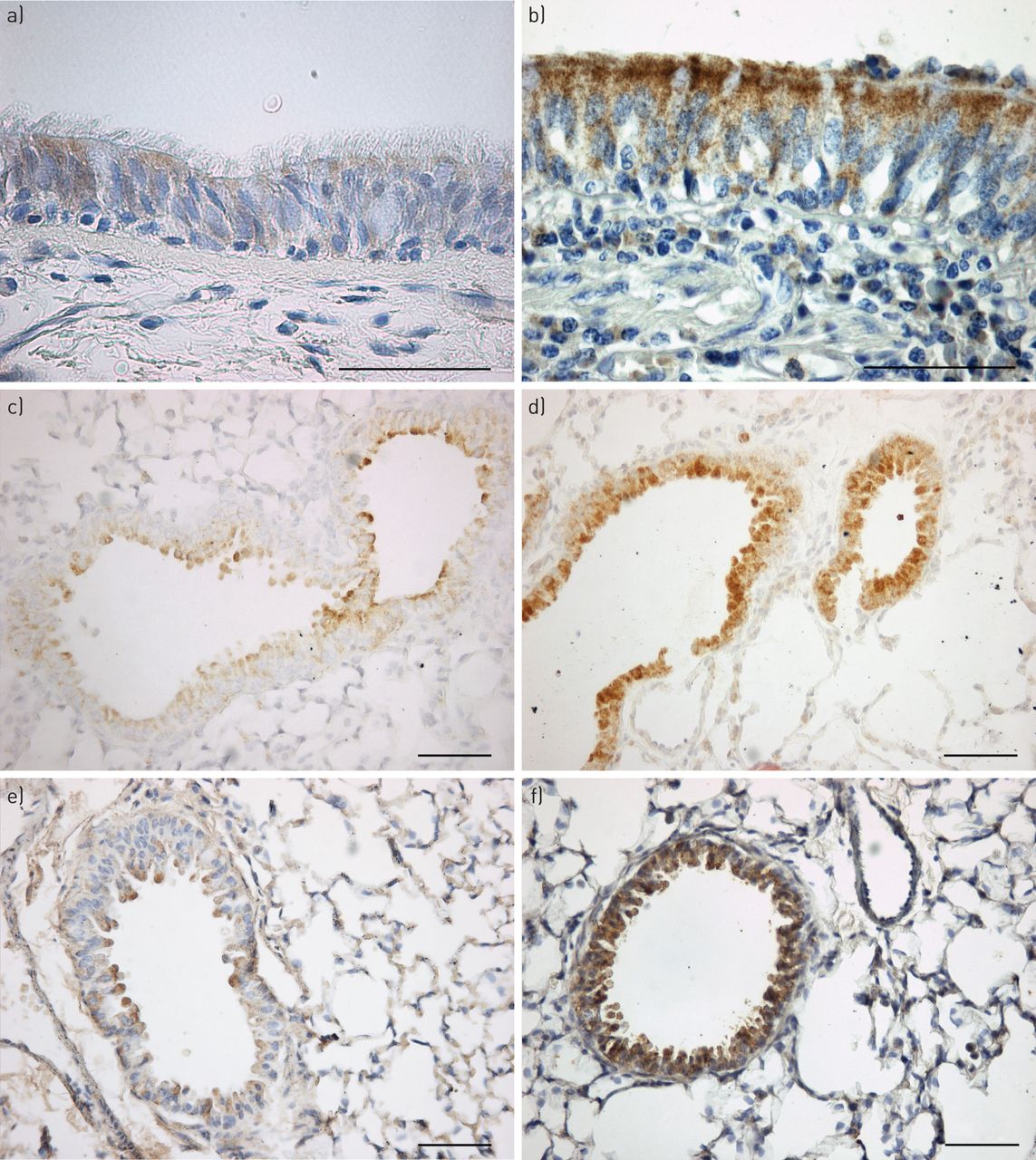
代表的疣状显微照片VEGF-A在人类和老鼠所示图3。代表显微照片的vWF疣状在人类主体在网上补充图S1。
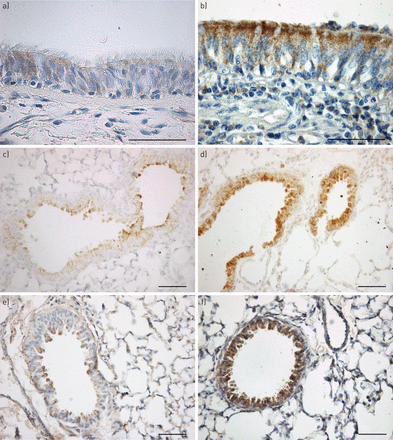
代表显微照片的血管内皮生长因子(VEGF)——在人类和鼠的气道免疫染色。气道部分得到的)人类控制的病人与b)囊性纤维化(CF)的病人,在c)囊性纤维化跨膜电导调节(雌性生殖道)+ / +与d)雌性生殖道- / -老鼠和e)F508wt / wt与f)F508▽/▽老鼠。部分被沾染了抗体VEGF-A(棕色),用苏木精复染色。在病人和控制雌性生殖道+ / +和F508wt / wt老鼠,只有稀疏的染色VEGF-A存在于上皮细胞。CF患者,在这两种雌性生殖道- / -和F508▽/▽老鼠,广泛VEGF-A存在于上皮细胞的染色。显微照片的结果代表10 10 CF患者,控制和7雌性生殖道+ / +和雌性生殖道- / -老鼠,在五个F508wt / wt和F508▽/▽老鼠。酒吧= 50μm规模。
雌性生殖道抑制效应在人工培养的气道上皮NCI-H292 VEGF-A合成细胞
短路电流测量证实NCI-H292细胞氯运输由CFTR选择性检测抑制剂,抑制CFTR-inh172年或ppq - 102(见在线补充图2)。治疗与CFTR-inh NCI-H292细胞172年(10−5米)或ppq - 102 (10−5米)诱导增加VEGF-A合成二(p < 0.05)与基线)(图4)。同样,NCI-H292细胞转染与特定的雌性生殖道siRNA VEGF-A合成增加(p < 0.05)与基线)(图4 b]),而转染不属预定目标的核没有效果。
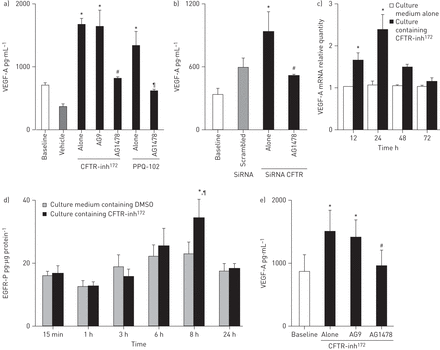
囊性纤维化跨膜电导调节效应(雌性生殖道)抑制血管血管生成生长因子(VEGF)——合成和表皮生长因子受体(EGFR)在气道上皮细胞活化。CFTR)效应的选择性检测抑制剂在NCI-H292 VEGF-A合成细胞:细胞培养在培养基(基线),培养基含有二甲亚砜(车辆)或包含选择性雌性生殖道抑制剂(CFTR-inh培养基172年或ppq - 102;10−5米)。VEGF-A浓度是衡量ELISA(见方法部分)。在一些实验中,细胞使用选择性表皮生长因子受体酪氨酸激酶抑制剂AG1478 (10−5米)或不活跃的模拟AG9 (10−5米)。结果表示为±扫描电镜n≥3独立重复实验。*:p < 0.05与基线;#:p < 0.05与CFTR-inh172年独自一人;¶:p < 0.05与ppq - 102。b)转染效果与雌性生殖道小干扰RNA (si)在NCI-H292 VEGF-A合成细胞。细胞培养仅在细胞培养基(基线),培养基转染与不属预定目标的小干扰rna或培养基(炒)与特定的雌性生殖道核(见方法部分)。结果表示为±扫描电镜,n≥3独立重复实验。*:p < 0.05与基线;#:p < 0.05与雌性生殖道siRNA孤单。CFTR-inh c)的影响172年在NCI-H292 VEGF-A mRNA表达细胞。细胞培养在培养基(基线)或包含CFTR-inh培养基172年(10−5米),VEGF mRNA量化使用定量PCR和报道的相对数量。n = 3独立重复实验。*:在每个时间点与基线相比p < 0.05。CFTR-inh d)的影响172年在NCI-H292细胞EGFR磷酸化。NCI-H292细胞培养在培养基含有二甲亚砜(DMSO)或包含CFTR-inh培养基172年(10−5米)。在不同的跨度为收集细胞溶解产物,和磷酸化表皮生长因子受体(EGFR-P)浓度测定ELISA(见方法部分)。结果表示为±扫描电镜从n≥3独立重复实验。*:p < 0.05与DMSO 8 h;¶:p < 0.01与CFTR-inh172年在15分钟CFTR-inh e)的影响172年在VEGF-A合成人工气道上皮细胞的主要文化(HAECs)气液界面。在21天的文化,HAECs仅在培养基培养(基线)或包含CFTR-inh培养基172年(10−572 h M)。结果表示为均值±SEM从n = 5独立重复实验。*:p < 0.05与基线;#:p < 0.05与CFTR-inh172年一个人。
检查是否雌性生殖道的作用抑制VEGF-A合成与转录效应,我们检查了使用qPCR VEGF-A mRNA的表达。治疗与CFTR-inh NCI-H292细胞172年(10−5米)诱导增加VEGF-A mRNA在12 h,达到24小时,然后(见减少图4摄氏度)。
的角色在VEGF-A合成雌性生殖道引起抑制EGFR磷酸化NCI-H292细胞
因为EGFR磷酸化VEGF-A合成,需要由各种刺激诱导气道上皮细胞(18,29日),我们检查它在VEGF-A合成雌性生殖道引起的抑制作用。治疗与CFTR-inh NCI-H292细胞172年诱导显著增加EGFR磷酸化8 h (p < 0.05与车辆,p < 0.05与CFTR-inh172年在15分钟)(图4 d)。与AG1478预处理NCI-H292细胞(10−5M,选择性表皮生长因子受体酪氨酸激酶抑制剂)阻止VEGF-A合成诱导雌性生殖道化学抑制剂或雌性生殖道核(p < 0.05)与基线)(图4 a和b)。与AG9预处理NCI-H292细胞(10−5米,一个不活动的模拟AG1478)没有影响。
讨论
我们发现,气道上皮的主要网站VEGF-A疣状在CF受试者获得肺移植,这VEGF-A表达式在CF与对照组相比增加了。VEGF-A疣也增加了在两个不同品系小鼠气道上皮细胞和雌性生殖道功能障碍。在人工气道上皮细胞,抑制雌性生殖道(使用化学抑制剂或siRNA)导致VEGF-A合成增加,所需的表皮生长因子受体激活。这些数据表明,雌性生殖道功能障碍导致VEGF-A upregulation CF气道上皮的科目。他们进一步建议角色上皮VEGF-A支气管旁血管重塑的发生在CF航空公司。
我们报告首次在CF VEGF-A气道上皮的表达增加。米cColleyet al。(12)报道,增加血清VEGF-A水平稳定的CF科目与主题与non-CF肺疾病,但VEGF-A水平没有测量气道的隔间。米注视者et al。(13]类似的浓度测量的VEGF-A支气管肺泡灌洗液的九个CF和16的同龄健康志愿者。因为VEGF-A退化在体外CF的支气管肺泡灌洗液,包含高中性粒细胞弹性蛋白酶活性,这些作者无法得出一个明确的结论对肺生产VEGF-A CF学科(13]。Krennet al。(15发现增加VEGF mRNA和蛋白在肺匀浆CF的主题在不吸烟者的肺移植而获得。后面我们的数据证实了这些发现和扩展这些数据表明,气道上皮的主要网站VEGF-A表达CF肺。
我们得出结论,雌性生殖道功能障碍引发VEGF-A合成是基于获得的数据在培养气道上皮细胞。此前,Verhaegheet al。(30.]报道增加VEGF-A合成在人类胎儿带着纯合突变细胞系F508del而控制人类胎儿细胞线。这些后者数据提供旁证的雌性生殖道的作用功能障碍VEGF-A生产;然而,这两个细胞系之间差异可能是与雌性生殖道功能障碍[无关31日]。在目前的研究中,使用化学抑制剂抑制雌性生殖道通道功能,或可拆卸的小干扰rna在雌性生殖道使用雌性生殖道non-CF细胞系和non-CF HAECs提供了直接证据,雌性生殖道功能障碍引发VEGF-A合成。增加了气道上皮的VEGF-A疣状雌性生殖道有缺陷的小鼠为这一发现提供了进一步的证据。
发现增加VEGF-A表达增加支气管旁血管供应无关雌性生殖道缺乏的老鼠有点奇怪。因为VEGF-A刺激内皮细胞增殖和血管生成通过其影响血管内皮生长因子受体2 (VEGFR2) [32),我们在小鼠肺检查VEGFR2的表达。野生型和雌性生殖道不足(包括雌性生殖道- / -和F508▽/▽VEGFR2)小鼠,疣状是局部的气道上皮细胞,但没有在支气管旁血管内皮(见在线补充图。S4)。相比之下,Baluket al。(11]发现VEGF-A超表达在气道上皮细胞,导致气管血管生成,与表达式的VEGFR2气管血管的内皮细胞。我们建议缺乏VEGFR2表达内皮可能占的缺席增加血管供应雌性生殖道有缺陷的老鼠。
我们的研究依赖于使用的力量互补模型和信息来源,包括人类CF组织、小鼠模型和培养HAECs。我们也认识的局限性的方法。之间的相关性发现VEGF-A上皮和支气管旁多血管显著只有当分组控制和CF科目。支气管旁血管之间缺乏相关性和上皮VEGF-A CF学科可能与CF组织可用的数量相对较少,并且这些患者招募的齐次特征移植。另外,上皮VEGF-A不得与CF的支气管旁血管生成相关主题和可能扮演不同的角色(如。上皮增生(33])。CF比对照组年轻受试者在移植。因为一项研究VEGF-A疣状在年轻人与正常肺功能报告没有疣状气道上皮细胞(34),这种差异不太可能影响我们的发现。小鼠模型的雌性生殖道功能障碍不繁殖人类CF肺部疾病的典型结构异常(35]。因此,没有增加支气管旁血管供应雌性生殖道有缺陷的小鼠应谨慎解释。然而,一个可能假设,除了雌性生殖道功能障碍,其他因素诱导VEGF-A表达式,如细菌感染(18),促进支气管旁血管生成可能是必要的。最后,雌性生殖道抑制诱导VEGF-A信使rna和蛋白质合成增加,表明转录效应。然而,进一步的研究将是必要的,以确定是否影响信使rna合成或mRNA稳定有关。
我们的研究结果表明,雌性生殖道功能障碍引发VEGF-A在气道上皮细胞合成,独立于感染和炎症。然而,增加VEGF-A表达式出现在CF气道上皮细胞在移植可能是与多种因素有关:促炎细胞因子(如。肿瘤坏死factor-αinterleukin-1β),从革兰氏阴性和革兰氏阳性细菌(微生物产品18,29日)和病毒(29日),和缺氧36)也被报道增加VEGF-A在气道上皮细胞合成。
VEGF-A本地化纤毛细胞,可能由于纤毛细胞表皮生长因子受体激活或纤毛细胞的前体(如。基底细胞)。其他主要产品在气道上皮细胞(表皮生长因子受体的激活如。黏蛋白和interleukin-8)一直在局部分泌细胞(4]。本地化的表皮生长因子受体在气道上皮细胞亚群有点争议的文学:Tynet al。(37]报道的选择性表达EGFR在纤毛细胞在活的有机体内在培养的上皮细胞,并建议纤毛细胞的分化转化成粘蛋白生产细胞。其他人发现表皮生长因子受体在基底细胞和nongranulated分泌细胞,但不是在纤毛细胞(4,38]。建议EGFR本地化是一个动态的过程,在不同病理生理条件下可能会有所不同。
使用CFTR-inh抑制雌性生殖道功能172年引发了表皮生长因子受体磷酸化,并增加VEGF合成所需的雌性生殖道抑制表皮生长因子受体激活后气道上皮细胞。治疗与CFTR-inh NCI-H292细胞172年导致抑制氯运输几分钟后,证实了先前的发现Perezet al。(26),但EGFR磷酸化增加8 h。因为激活表皮生长因子受体的配体(如。表皮生长因子(EGF)或改变增长factor-α)通常发生在几秒钟之内,这些数据表明CFTR-inh172年不是一个表皮生长因子受体配体,表明激活表皮生长因子受体在这种情况下发生通过一连串的事件在上皮细胞。的精确分子机制导致表皮生长因子受体激活仍有待建立。
VEGF-A合成增加气道上皮细胞导致血管生成(11,30.)和增加血管通透性,促进招聘炎症细胞(如。中性粒细胞)和蛋白质的航空公司。此外,支气管旁血管生成可能导致发生大咯血,CF患者的潜在威胁生命的并发症(39]。大量咯血CF患者通常是治疗支气管动脉embolisation。高复发后观察embolisation在这些患者中,可能与持续的支气管旁血管增生和炎症有关。反复咯血控制通过反复embolisation CF患者的肺移植是一个公认的标准。我们建议直接针对血管新生疗法在CF反复咯血患者可能是有用的,但还需要进一步的研究在这种疗法可能被认为是人类。
总之,VEGF-A表达显著增加在CF气道上皮移植,显示角色在CF患者支气管旁血管生成(11]。雌性生殖道功能障碍直接导致VEGF-A upregulation在气道上皮细胞通过表皮生长因子受体激活。治疗直接针对雌性生殖道功能障碍(40,41)可能导致VEGF-A合成和支气管旁血管的显著减少。此外,生长因子(如。EGF和/或VEGF)受体拮抗剂可以代表一个有前途的方法减少血管重塑CF航空公司。
确认
作者感谢索菲Danel-Beaucaire(科钦医院,法国巴黎)免疫组织化学染色和克莱尔Danel (Bichat医院、巴黎、法国)提供人类CF组织。
脚注
社论评论看1438页。
可以从本文的补充材料www.www.qdcxjkg.com
支持声明:这项工作是由赠款Chancellerie des巴黎大学(腿Poix),协会Vaincre la Mucoviscidose和协会Cardif。
利益冲突:披露可以找到与本文的在线版本www.www.qdcxjkg.com
- 收到了2012年10月14日。
- 接受2013年3月5日。
- ©2013人队











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}