条文本

文摘
的蛋白质和分子特征变异的α1抗胰蛋白酶(AAT)基因,AAT的遗传流行病学和可用的数据不足。
- α1-antitrypsin不足
- 基因型
- 等电点聚焦
- 人口筛查
来自Altmetric.com的统计
在过去的40年,出版后Laurell和埃里克森,开创性的论文1有重大的进步的理解基因异常与α有关1抗胰蛋白酶(AAT)缺乏症和病理生理学的肺和肝脏疾病相关。在同一时期,许多遗传流行病学调查数据积累。结果,我们现在有一个相当全面的照片AAT缺陷的分布,特别是在发达国家,和一些良好的基础假设AAT基因进化,缺AAT的起源,它的传播。本文综述了缺AAT的遗传流行病学数据可用。初步讨论AAT变异的蛋白质和分子特征提供一个背景,以促进更好的理解所讨论的术语和流行病学数据。
AAT蛋白
α1抗胰蛋白酶(AAT或α1——也称为α1蛋白酶或蛋白酶抑制剂(α1-π))是一个52 kD糖蛋白主要由肝细胞分泌,在较小程度上,通过肺上皮细胞和吞噬细胞。它能抑制多种丝氨酸蛋白酶但它的首选目标是人类嗜中性粒细胞弹性蛋白酶(HNE),它演示了协会最高速率常数。2AAT在肺部的主要功能是保护HNE释放引起中性粒细胞的结缔组织,所支持的对象在生命的早期肺气肿的发展受到严重影响AAT的遗传缺陷。3在大多数人类肺部辩护从HNE攻击正常AAT等离子体水平从100到200 mg / dl(以浊度测定法)。尽管AAT是一个众所周知的急性期反应物,其变异性大正常血浆水平主要反映了多形性的糖蛋白。超过100的遗传变异的AAT已确定这些是严格与特定AAT等离子体水平共显性遗传方式4,5——换句话说,等离子体水平都由AAT基因等位基因彼此独立的。
目前使用的术语识别AAT变异是一种妥协的演变所产生的不同的技术应用于蛋白质分离和描述在过去的40年。AAT变体包括在一个等位基因的系统称为π(蛋白酶抑制剂)系统最初命名的基础上他们的迁移速度在米淀粉凝胶电泳(媒介),S(缓慢),F(快),或Z(很慢)。6随后,当蛋白质开始被分离的基础上他们的等电点(pH值4 - 5在薄层聚丙烯酰胺凝胶等电聚焦()),以应对前面的命名系统AAT变异与字母表的第一个字母分类如果显示阳极的迁移和最后一个字母显示阴极的迁移。在基因组时代的到来前π体系改名为π*识别AAT基因位点。5
Laurell原始论文和埃里克森之后1和随后的证据表明,大多数科目继承严重缺乏AAT倾向于肺气肿的早期发病,7它成为有用的临床用途分类AAT变异分为三大类:4
正常的,特点是AAT等离子体水平一般人群的参考范围之内,与肺或肝脏疾病的风险。这个类别包括四个最常见的中间迁移M变体(M1→M4)和一些不太常见的变异识别的基础上字母字母,如上所示,和这座城市最古老的生活载体的变体8——例如,L法兰克福。
不足的特点是减少,但检测AAT等离子体水平,增加患肺癌的风险或肝脏疾病。这一类包括最常见的有缺陷的变异、Z和年代,许多少变体包括所谓M-like变体(M马尔,米普罗奇达等),中间迁移模式。等离子体的上限水平在这个类别包括AAT变体是80毫克/ dl-that,显示大多数科目的π*深圳基因型。
零(目前问:指定),没有检测到等离子体水平,增加患肺气肿的风险。
AAT基因及其演化
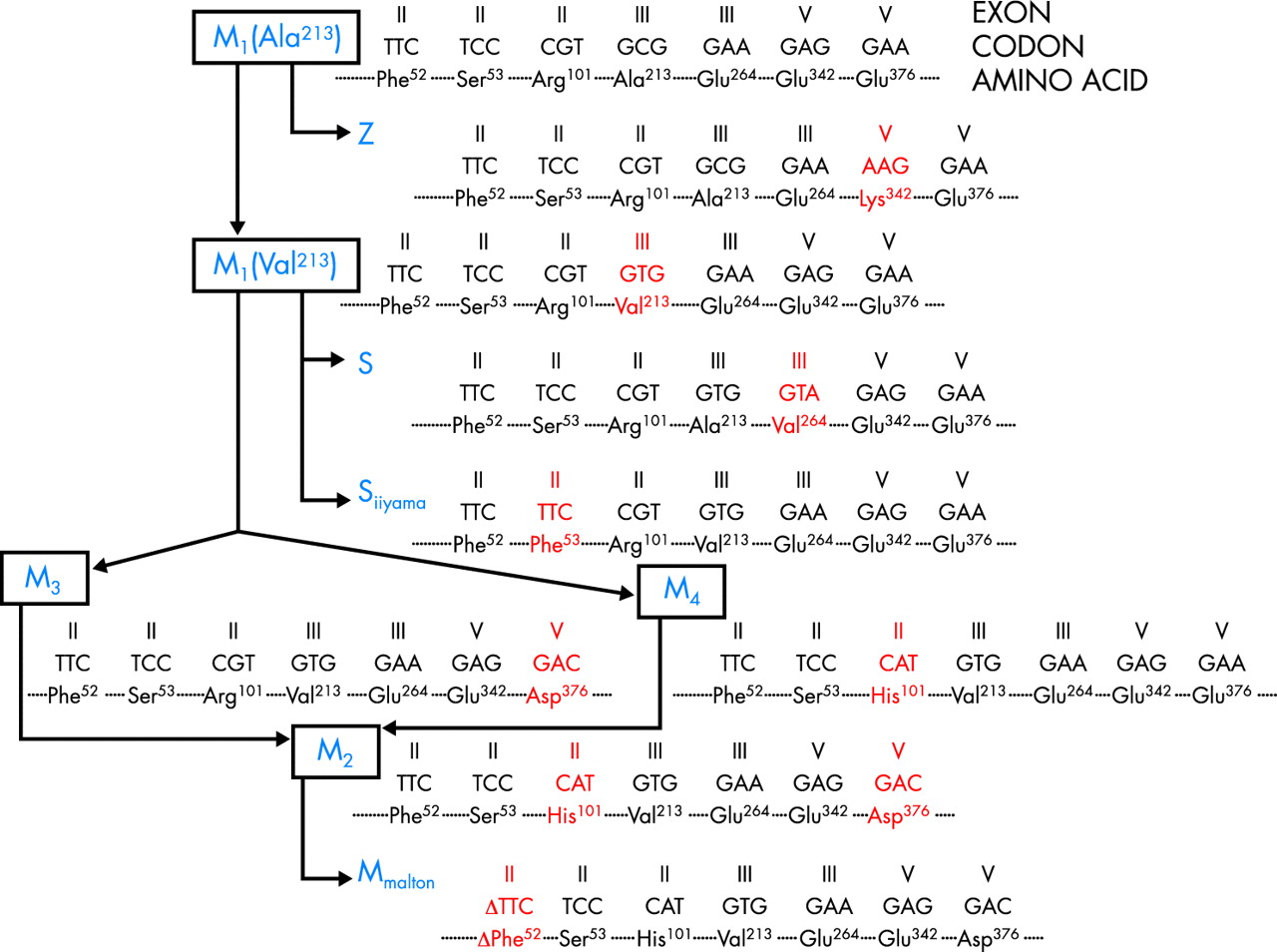
AAT基因的分子结构的知识开始出现大约二十年后报告的血清蛋白缺乏。9AAT基因是基因簇的一部分,位于人类染色体14 q32.1,称为SERPIN (SER快乐Proteinase在hibitor)表生的。这个基因集群包括皮质类固醇激素结合球蛋白(cbre), AAT-like假基因(公益诉讼),AAT,蛋白C抑制剂(PCI)和α1-antichymotrypsin (AACT)基因(着丝点染色体端粒)。AAT横跨12.2 kb的长度和有三个非编码基因(IA、IB、IC)和四个编码(II, III, IV, V)外显子;外显子V包含序列编码AAT蛋白的活性网站(满足358年硒359年)。之间有一个紧密的遗传连锁AAT和AACT基因,这可能是这两个位点分化相对最近(100 - 250年前)。10只有几个核苷酸之间的差异已发现AAT在高等灵长类动物(狒狒、大猩猩和黑猩猩)和人类AAT祖传的变体,所有显示参数101年阿拉巴马州213年-Glu376年组合。11基于这三个主要的氨基酸的替换单体型标记,它已经可以画一个可能主要的正常AAT变异和种系发生树最经常发现缺陷的变异(图1)。
{kind=link}

{kind=link}
AAT基因的系统发育树。修改从Nukiwa等。11
研究AAT缺陷的分布
虽然,正如下面所讨论的,大量的人群进行了调查,只有少数人口研究基于AAT等位基因频率,主要基于献血者筛查。两个开创性的论文最近回顾可用的数据缺AAT的地理分布。12日,13鉴于欧洲广泛接受障碍出现人口,和记黄埔是逻辑来解决他的论文在AAT缺陷的分布在欧洲。12然而,在日益增长的意识之光遗传条件,de serre13最近扩大了前面分析的总结全球调查,还看种族/民族缺AAT的患病率的差异。都评论了出版遗传流行病学调查和大量的研究(373年德serre控制军团”13),这样,第一次,我们有一个global-if没有AAT缺乏全面的看法的。当然,评论都有固有的局限性荟萃分析的研究,在许多方面都有所不同。12
选择组
调查包括各种各样的主题:献血者,新生儿,孕妇、各种组织的工人,学生,和主题提交血统测试,实验室或医院工作人员,或选择“随机”个体。在许多情况下没有选择标准的详细信息是可用的。
样本大小
调查样本大小从几十到几千不等,有很高比例的调查包括100 - 500年的主题。基因频率估计群不到200个人有一个错误的风险很高。
的方法确定AAT的变体
在大多数的研究能源论坛AAT变异是由合作(也称为“表现型”),尽管交叉免疫电泳是用在许多老一辈的调查发表在1980年代。这两种方法并不完全可以互换。5到目前为止,没有遗传流行病学调查AAT缺乏使用分子方法(pcr)的报告。考虑到这些差异,一定程度的偏差可能被引入到全球的分析调查。
遗传流行病学调查分析
三个主要的流行AAT变体(π* M,π* Z,和π* S)在大多数调查报告基因频率,也就是该等位变异的频率(变异的贡献两个等位基因)或在杂合子(贡献一个等位基因的变体),和援引0.0……n(或n每1000人)。超越是使用基因频率与哈迪温伯格平衡公式来估计运营商的总数(π*女士和π* MZ)和主体缺乏变体组合(π* SS,π*深圳,和π* ZZ)。这种方法是由德serre使用13估计人口风险与缺陷变异(运营商+主题)在一个特定国家或地理区域,考虑到整个人口。
遗传流行病学AAT的缺乏
欧洲
π* Z患病率最高的变体被记录在欧洲北部和西部国家基因频率(平均0.0140),13峰值在斯堪的纳维亚半岛南部、丹麦、荷兰、英国和法国北部(基因频率> 0.0200)。14 -20.结果整个人口的大规模筛查新生儿在瑞典进行2年时间于1976年出版。21129 200 000婴幼儿,π* Z变异,产生频率为1 1550年个人和基因频率为0.026。Sveger也筛选11 18岁000名身体健康的男性,发现5π* Z和10π*深圳个人。22最近发表的研究主题是来自哥本哈根城市心脏研究中随机选取的9187名被试进行调查。23本研究中的患病率在1500年(1个人)是一样的,在瑞典的研究中,21但是π* Z基因频率是0.049。π* Z的患病率逐渐减少在欧洲国家在西北→东南方向,最低的数据被记录在东欧。12
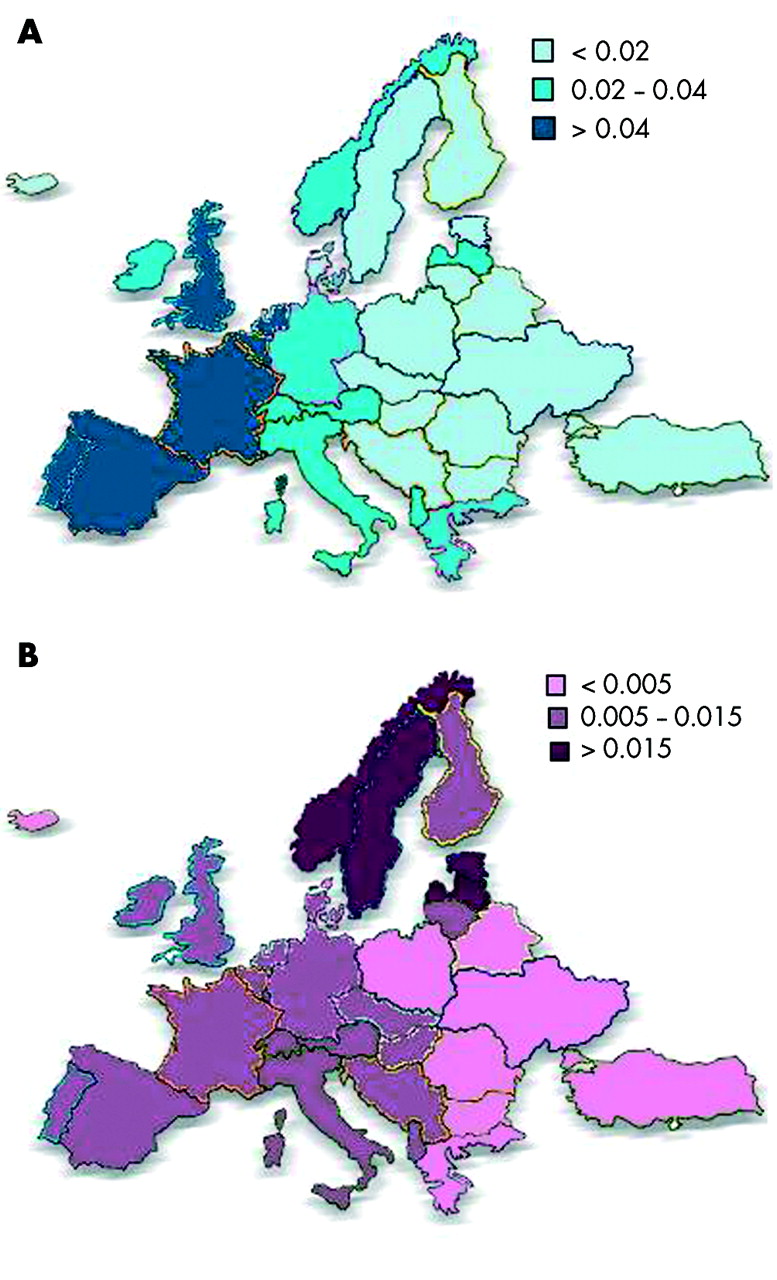
π* S的分布明显不同于π* Z和更均匀。24π* S的最高频率是在南欧基因频率(平均0.0564),13峰值在伊比利亚半岛(基因频率> 0.1400)。25日,26π* S的分布沿着→东北西南梯度逐渐减小。的分布π* Z和π* S在欧洲列于图2。
{kind=link}

{kind=link}
频率(A)π*年代和(B)π* Z基因在欧洲。
π* S的平均比率:π* Z是4.5:1在南欧,西欧的3.5:1、1.1:1在北欧(从德serre计算等13)。
遗传流行病学的AAT不足尤其是欧洲人
如上所述,斯堪的纳维亚半岛是欧洲最高的地区之一为π* Z型数据;然而,基因频率对π* S和π* Z在芬兰和瑞典拉普人在欧洲最低的频率。27日,28
分析40从意大利军团(de serre审查通过的等29日)显示,基因频率的π*年代和π* Z是最高的在意大利北部和减少逐渐从北到南。在为数不多的人口基础调查执行9000年新生儿在南蒂罗尔,比较德国和意大利人产生π* Z基因频率为0.019和0.015,分别。30.在撒丁岛人π* S的基因频率高于意大利大陆而π* Z是低得多。
π* S在巴斯克地区的频率高达在伊比利亚半岛的其他部分,而π* Z型的频率低得多。31日,32
尽管可以假设,至少在拉普人,等肺部疾病的易感基因π*年代和π* Z可以消除不利的气候,更有可能的解释的多样性π*基因频率位于拉普兰人的孤立,撒丁岛人,巴斯克种群与其他基因的影响。事实上,分析主要组织相容性复合体类我等位基因显示,这些人群有显著的遗传差异与其他周围的人群。33
其他发达国家
北美
以来人们普遍认为AAT缺陷出现在欧洲人群的传播障碍在欧洲国家的居民有一个背景并不奇怪。34然而,π* Z的平均基因频率在北美是0.0092(在最低范围报告结束在欧洲),而π* S基因的频率为0.0328,高于欧洲北部的报道。13这个数字可能是由于混合种群在北美和可用的有限数量的人群(43)对整个人口。基于人口抽样调查的执行000年20献血者在圣路易斯地区产生了一个π* 2800年Z的1个人。35
澳大利亚和新西兰
基因频率的π* Z和π* S在澳大利亚和新西兰非常接近那些报道北美(分别为0.0151和0.0395),可能出于同样的原因。
远东亚洲
非常有限的军团已报告来自日本、中国和韩国。13π* Z是0.0002的基因频率在日本,在中国0,和0.0061在韩国,而各自的π* S数据是0.0004,0.0006和0.0070。有趣的是,AAT M1(阿拉巴马州213年)变体,发现大约有20 - 23%的AAT缺乏白色主题,11没有检测到任何的156名日本主题。自开发了M1 (Ala Z变体213年)基地等位基因(图1),这也许可以解释极端罕见的π* Z远东日本和其他人群的基因。这些发现也占了年代的流行iiyama日本缺乏变体;这种变体出现的M1 (Val 213)基地等位基因(图1)36岁,37并在156年日本调查的100%。
南美
只有少数群体从美国南部已报告,所以没有公司可以提供数据。13
发展中国家
相信AAT缺乏症是一种障碍,主要影响白色主题,在一定程度上动摇了全球调查的分析由de serre。13他提供的证据相当流行的人群中π* Z和π* S从中东和北非,中部和南部非洲、中亚和东南亚地区,表明缺AAT已经胜过了种族和民族的界限。
年龄估计AAT缺乏变异及其在人群中的扩散
分析serpin内的等位变异基因簇定义人群可能产生有用的信息关于起源的时间和地点AAT缺乏变体。在调查白色π* Z家庭起源于北欧,深海角和同事38发现,97%的病例有独特的60 kB包括cbre单体型,公益诉讼,和AAT与π* Z相关的基因等位基因,从而支持单一起源的理论π* Z突变。39单体型分析还允许估计第一π* Z突变发生的时间。基于的假设在给定地区随机重组,深海角提出π* Z突变可能出现66代前,假设33年的平均延迟每一代,~ 2000年前。这估计不同于前一个假设的216代(~ 7000年前39)和最近先进估计的120代(~ 4000年前40)。有趣的是,根据去年估计,π* Z突变可能是分散在新石器时代,为囊性纤维化已经建议ΔF508突变。40岁,41根据假设在一个给定的国家基因频率越高,越有可能是,基因首先发生,12普遍接受的是π* Z基因出现在北欧(斯堪的纳维亚南部也许更准确),随后蔓延到欧洲其他国家的地中海国家,后被北欧海盗航行等主要欧洲人口流动。然而,单体型的事实模式多样性与预期的逐步减少,如果突变传播从北到南,从伊比利亚半岛在人群,40和π* Z类型被发现在人群从中部和南部非洲和亚洲,13建议对人口流动的主要已知的方向扩散,或者或者,multiregionalπ* Z基因起源。
的发现π* S的患病率最高的是伊比利亚半岛表明π* S基因可能起源于这个领域,或者更确切地说在葡萄牙人口。40有趣的是,serpin单体型该人群的调查表明,这一事件发生15 000 - 000年(450 - 300代)前,使π* S突变比π* Z。在欧洲的传播π* Z,π* S的西向东梯度突变表明扩散对已知的主要在欧洲人口流动。
全球估计缺AAT的主题
考虑基因频率的π*年代和π* Z遗传流行病学调查报告的国际文献的检索和总人群中的个体的数量在不同的国家,de serre13全球计算估计的影响中间AAT不足的科目(运营商)和高患肺癌的风险/肝病与AAT不足(也就是说,π* Z比如和π*深圳复合杂合的)。当然,这种方法具有潜在的偏见:除了上述局限性的研究报道认为,一些调查进行选择的人群中很差一般人群的代表。29日为例,对意大利军团检查包括许多峡谷南边的阿尔卑斯山30.(地质屏障很明显,分离这些军团从这些居民中招募了附近的平原),和撒丁岛(其遗传隔离已经进行了讨论33)。这些结果应该谨慎考虑。尽管有这些警告,总体估计大约116 000 000运营商和1 100 000例严重缺AAT全球令人震惊和表明AAT不足可能是最常见的一种严重遗传性疾病(表1)。
开放式问题和未来的发展方向
AAT尚未得到充分认识不足是一个高昂的状态
考虑上述估计,只是明显可见,医生积极参与诊断和管理AAT的缺乏症,而这是一个尚未得到充分认识很大程度上高昂的状态。AAT替代疗法对个人的可用性肺气肿与AAT不足有关42鼓励科学界建立和加强AAT缺乏筛查项目在发达国家,即使在那些没有以往被视为高发病率的疾病,并实施国家注册。43在回应一个建议世卫组织会议期间转发AAT不足,34几个国家的国际注册邦联国家注册成立于1996年。44岁的45尽管广泛的努力来识别在AAT缺陷的情况下,很明显,只有少数的主题实际上是承认(表2)。
这under-recognition至少有两个原因。首先,相关的临床表型AAT不足(肺气肿、慢性支气管炎、支气管扩张、哮喘,并在较小程度上,慢性肝病)并非唯一条件。甚至表型的家族聚集性,遗传疾病的一个典型特征,并不是一个有用的信号从常见的慢性阻塞性肺疾病往往倾向于集中在家庭。46其次,π* Z基因的特点是一个不完整的penetrance-that,基因型与临床表型的关系不强。西尔弗曼和同事检查肺功能在一群52π* Z主题:20的52个科目(38%)在1秒用力呼气量(FEV1)超过65%的预测,经常在正常范围内。47这些π* Z的主题与正常或轻度肺功能受损通常确认为非索引病例,病例在家庭筛选确定。相同的作者还发现,疾病的严重程度表现受到一些变量,如吸烟和下呼吸道感染(×基因环境交互作用)。对象与无症状或轻微缺AAT的比例甚至更高一系列94人中π*深圳复合杂合性。48总之,有证据表明,许多人患有严重缺AAT没有临床意义上的肺功能障碍。这个特性缺乏AAT值得进一步调查,从流行病学和遗传的观点。
流行病学罕见的(非z non-S) AAT缺乏变体
对罕见的遗传流行病学AAT缺乏变异被认为是不超过2 - 4%的变异。49然而,这些变异可能的患病率高于此前认为因为罕见AAT缺陷变异可以被误认为是π* Z变体,因此误诊。事实上,我们从意大利注册AAT初步数据缺乏表明,多达22%的总AAT缺乏变体是罕见的。50其中一些变异的命名法(见上图)反映了他们可能意大利起源(M普罗奇达,米巴勒莫,问:普罗奇达伊索拉迪,问:trastevere)。5一个有趣的问题是:是罕见AAT缺乏变异更频繁的在这些国家的基因频率较低的是这款?撒丁岛的数据似乎支持这一假设。51临床表型与常见AAT缺乏变体π* Z相当明确,稍后将讨论综述系列,但是目前没有信息可在临床表型与罕见AAT缺陷变异有关。这应该是解决未来的研究。
确认
作者感谢Drs乔丹Baccheschi Ilaria Ferrarotti,和米歇尔Zorzetto帮助在准备这个手稿,雷切尔博士和sten编辑它。
引用
脚注
部分支持IRCCS亲自到圣马特奥Ricerca Corrente赠款、意大利MIUR Progetti di感兴趣重回2002年,基金会Cariplo,拜耳欧盟,Altana制药公司。