条文本

摘要
背景慢性阻塞性肺疾病(COPD)的发病机制尚不清楚。microrna (microrna或microrna大鹏展翅)是调节特定基因和蛋白质水平的非编码RNA小分子。确定mirna在COPD中的表达模式可以增强我们对疾病机制的理解。进行了一项研究,以确定mirna在患有和不患有COPD的吸烟者的肺部是否存在差异表达。比较miRNA和mRNA的表达,丰富与COPD发病机制相关的生物网络。
方法无阻塞性肺疾病证据的吸烟者(n=9)和患有COPD的吸烟者(n=26)的肺组织在验证后检测miRNA和mRNA的表达。然后,我们检测了miRNA和mRNA的表达,以丰富相关的生物学途径。
结果70个mirna和2667个mrna在COPD患者和非COPD吸烟者的肺组织中表达差异。miRNA和mRNA表达谱丰富,可能与COPD发病机制相关的生物学途径包括转化生长因子β、Wnt和局灶黏附途径。mir - 223和mir - 1274 a与没有梗阻的吸烟者相比,COPD受试者中受影响最大的mirna。miR-15b与没有梗阻的吸烟者相比,慢性阻塞性肺病样本中肺气肿和纤维化两个区域都有局限性。miR-15b在GOLD类COPD患者中表达差异。SMAD7的表达,被证实为miR-15b慢性阻塞性肺病患者支气管上皮细胞中。
结论与无梗阻的吸烟者相比,COPD患者miRNA和mRNA的表达存在差异。研究这些关系可能会进一步加深我们对疾病机制的理解。
- 慢性阻塞性肺疾病
- 肺气肿
- 微
- 基因表达
- COPD机制:COPD加重
- 慢性阻塞性肺病急性加重
- 慢性阻塞性肺病的病理
- 慢性阻塞性肺病药理学
- 肺气肿
- 肺减容手术
- 肺康复
- 烟草与肺
- 间质纤维化
- 慢性阻塞性肺病流行病学
- 肺癌
- 气道上皮细胞
来自Altmetric.com的统计数据
- 慢性阻塞性肺疾病
- 肺气肿
- 微
- 基因表达
- COPD机制:COPD加重
- 慢性阻塞性肺病急性加重
- 慢性阻塞性肺病的病理
- 慢性阻塞性肺病药理学
- 肺气肿
- 肺减容手术
- 肺康复
- 烟草与肺
- 间质纤维化
- 慢性阻塞性肺病流行病学
- 肺癌
- 气道上皮细胞
关键信息
关键问题是什么?
慢性阻塞性肺疾病(COPD)是一种主要由吸烟引起的全球性健康危机。
COPD发展的确切机制尚不清楚,但基因组学和蛋白质组学研究正在提高我们对这种疾病发病机制的理解。
研究人员已经在几种类型的癌症中发现了microRNAs (miRNAs或miRs)的异常表达,但我们对这些小分子在包括COPD在内的癌症以外的疾病中的重要性的认识才刚刚出现。
底线是什么?为什么要继续读下去?
在没有COPD的吸烟者和COPD患者(GOLD期1、2和4)的肺组织中发现了70个差异表达的mirna。
差异表达的mirna和基因之间的相关性提示可能与COPD发病机制有关的潜在功能关系。
介绍
慢性阻塞性肺疾病(COPD)是一种世界性流行病,主要可归因于吸烟的影响。它是一种不完全可逆的气流限制和肺部对有害刺激的异常炎症反应所定义的异质性疾病。1所涉及的机制包括那些直接归因于吸烟影响的机制,如细胞损伤、炎症和异常修复。2
小分子核糖核酸(microrna米尔)是一个小的非编码rna家族的成员(大约21-25个核苷酸长),编码在从动物和植物到病毒的生物体基因组中。3.这些分子代表了人类基因组中以前被认为是无功能的成分。mirna是关键生物学功能的组成部分,通过破坏转录物的稳定性或抑制蛋白质翻译来调节基因和蛋白质水平。鉴于miRNAs与其靶mRNA之间互补序列的相对冗余性,单个miRNAs具有同时调控数十至数百个基因的能力。事实上,据估计,mirna可能靶向多达三分之一的转录组。4
miRNA失调与多种疾病的发病机制有关,包括血液病和实体恶性肿瘤。miRNA在原发性肿瘤组织、血液、痰和尿液中的表达模式正在被研究作为疾病诊断和预后的生物标志物。这些miRNA标记可能最终应用于临床实践。我们对miRNAs在肺部疾病发病机制中的作用的了解正在迅速扩大,miRNAs似乎在维持肺部发育过程中的内稳态方面很重要,并且可能在肺部疾病中起致病作用。56此外,在动物模型中,mirna可能参与对外源刺激下肺炎症的调节。78
关于mirna在COPD中的作用知之甚少。两项研究调查了香烟烟雾对miRNA表达的影响。910在第一项研究中,暴露于烟雾中的大鼠和假手术组之间有24种mirna显著下调。9在第二项研究中,研究了烟雾暴露对小鼠miRNA表达的影响。10在这项研究中,大多数去调控的mirna也被下调。
类似的研究正在吸烟者和COPD患者的特定细胞类型中进行。Schembri等比较吸烟者和从不吸烟者气道上皮细胞中miRNA和mRNA的表达。1128个mirna差异表达,大多数在吸烟者中下调。佐藤等检查的表达mir - 146 a在COPD患者和非COPD患者的成纤维细胞中。12mir - 146 a据报道,通过呼气气流评估,培养成纤维细胞中的表达与COPD严重程度相关。
我们在这项研究中的目的是确定miRNA在COPD患者肺部是否存在差异表达,以及miRNA表达是否可能与mRNA表达相关,从而与COPD发病机制相关的生物学途径有关。我们比较了全球阻塞性肺疾病倡议(GOLD)不同阶段COPD受试者和无气流限制的吸烟者肺组织中miRNA和mRNA的表达模式。我们富集了我们那组去调控mrna中预测的miRNA靶标。最后,我们确定了转化生长因子(TGF)-β, Wnt和局灶粘附途径作为mirna可能与COPD发病机制相关的潜在途径。
方法
方法的其他细节显示在在线补充。
主题的选择
35个组织样本通过肺组织研究联盟批准的项目(#07-99-0008)获得,用于miRNA和mRNA分析。研究对象的临床信息包括年龄、性别、身高、体重和吸烟史。测试结果包括肺活量测定、病理诊断和圣乔治呼吸问卷健康调查的结果。
组织处理和RNA分离
使用冷冻断裂方案从冷冻组织样本中分离总RNA,然后在- 20°C下提取Trizol并沉淀过夜以增加小RNA的产量。RNA完整性在Agilent 2100生物分析仪上通过毛细管电泳检测。只有完整性数≥7的RNA被用于分析研究。
microrna的微阵列
根据制造商的说明(Exiqon, Woburn, Massachusetts, USA),使用Exiqon miRNA微阵列对COPD受试者(N=19)和正常吸烟者(N=8)进行miRNA谱分析。将标记的miRNA探针与miRCURY LNA阵列v.11.0在56°C下杂交16 h。杂交后,清洗阵列并使用ScanArray Express (Perkin Elmer, Waltham, Massachusetts, USA)以5 μm分辨率进行扫描。
信使RNA微阵列
使用Agilent Quick Amp Labeling技术(Santa Clara, California, USA)制备样品进行mRNA微阵列分析。然后将混合的载玻片清洗并用ScanArray Express (Perkin Elmer)扫描。
mRNA和miRNA微阵列数据处理
对于mRNA数据,所有样本的原始强度被合并,使用分位数方法归一化并转换为对数2规模。13选择所有样本的平均强度大于全球平均强度的现有探针,并用于进一步的统计分析。LIMMA14和QVALUE15使用R包对对照吸烟者和COPD样本之间mRNA和miRNA表达差异进行统计检验,并分别计算阳性假发现率(pFDR)。采用现有探针进行统计检验,增加真阳性与假阳性的比例;16差异表达的mrna和mirna定义为pFDR <0.05,组间变化至少±1.5倍。
定量反转录PCR验证
采用定量反转录PCR (qRT-PCR)对所有患者样本进行独立检测,检测个体miRNA (miR-15b,mir - 223,mir - 1274 a和mir - 424)和mRNA(端粒酶相关蛋白1 (TEP1)、白细胞介素6 (IL-6)、过氧化氢酶(CAT)和母亲抗十肢瘫痪同源物7(果蝇)(SMAD7)) (Applied Biosystems, Foster City, California, USA)。所有RT-PCR实验均为三个独立实验,对所有研究样本进行三次重复。根据2的计算,相对于mRNA的18s或miRNA的U43给出数据(−ΔCt).通过Student t检验或方差分析(见miRNA和mRNA探针序列在线补充表E1和E2), p<0.05为统计学显著性。
功能富集研究
为了鉴定京都基因与基因组百科全书(KEGG)17与一组基因相关的途径,我们使用DAVID18并获得了富集通路的列表。它们的p值是通过一种被称为EASE评分法的改进Fisher精确检验来计算的。18
目标预测和网络分析
使用TargetScan v5.1、PicTar和miRanda获得miRNAs的预测靶基因。对于差异表达的mirna,使用所有人类基因中的预测靶点和与差异表达基因重叠的预测靶点进行KEGG通路富集分析。为了描述KEGG通路中差异表达基因与miRNA之间的潜在关系,TargetScan预测的表达负相关(< - 0.5)的miRNA基因对被纳入通路图。使用Cytoscape生成修改后的KEGG通路图。19
原位杂交和共定位
SMAD7、细胞角蛋白AE 1/3和miR-15b按照前面的描述进行表达。20.开发后,用Nuance系统(剑桥研究与仪器,霍普金顿,MA)分析幻灯片。Nuance系统将蓝色和棕色信号分别转换为荧光蓝色和红色,然后将它们混合以确定一个给定的细胞是否包含这两种目标。阴性对照是不使用探针和使用乱置探针,如前所述。20.
转染研究和western blot分析
转染研究和western blot分析的详细信息显示在在线补充。
TGFβ治疗
如上所述,Beas2B细胞转染了重组或pre-miR15b。转染72 h后,细胞饥饿6 h,然后用TGFβ (5 ng/ml;Sigma,圣路易斯,密苏里州,美国)。在10、15和30分钟收获细胞,并评估磷酸化SMAD 3蛋白的表达(Cell Signaling, Danvers, Massachusetts, USA)。
迁移分析
使用IBIDI培养插入物(德国m
结果
研究对象
组织样本取自GOLD期1、2和4 COPD患者(N=26)和来自肺组织研究联盟的无阻塞证据的吸烟者(N=9)。在COPD或对照组织标本中没有病理或临床癌症诊断。GOLD 4期组织取自外植肺。无气流阻塞的吸烟者组织和GOLD 1期和2期组织主要通过楔形切除获得,病理未显示癌症证据。人口统计资料载于表1附带每个组的汇总统计数据。
所有受试者的平均年龄之间存在总体差异(p<0.0001)。GOLD 4期COPD患者比GOLD 1期COPD患者(p<0.001)、GOLD 2期COPD患者(p<0.001)和无气流限制的吸烟者(p=0.013)年轻(见在线补充表E6)。两组之间的包龄没有总体差异(p=0.106)。GOLD阶段4的受试者比没有气流限制的吸烟者有更多的烟雾暴露(p=0.068;见在线增刊表E7)。在St George呼吸问卷得分和预测的1 s百分比用力呼气量的总体差异如预期的那样显著(p<0.0001)(见在线补充表E8)。
microrna在COPD和对照肺组织中的表达存在差异
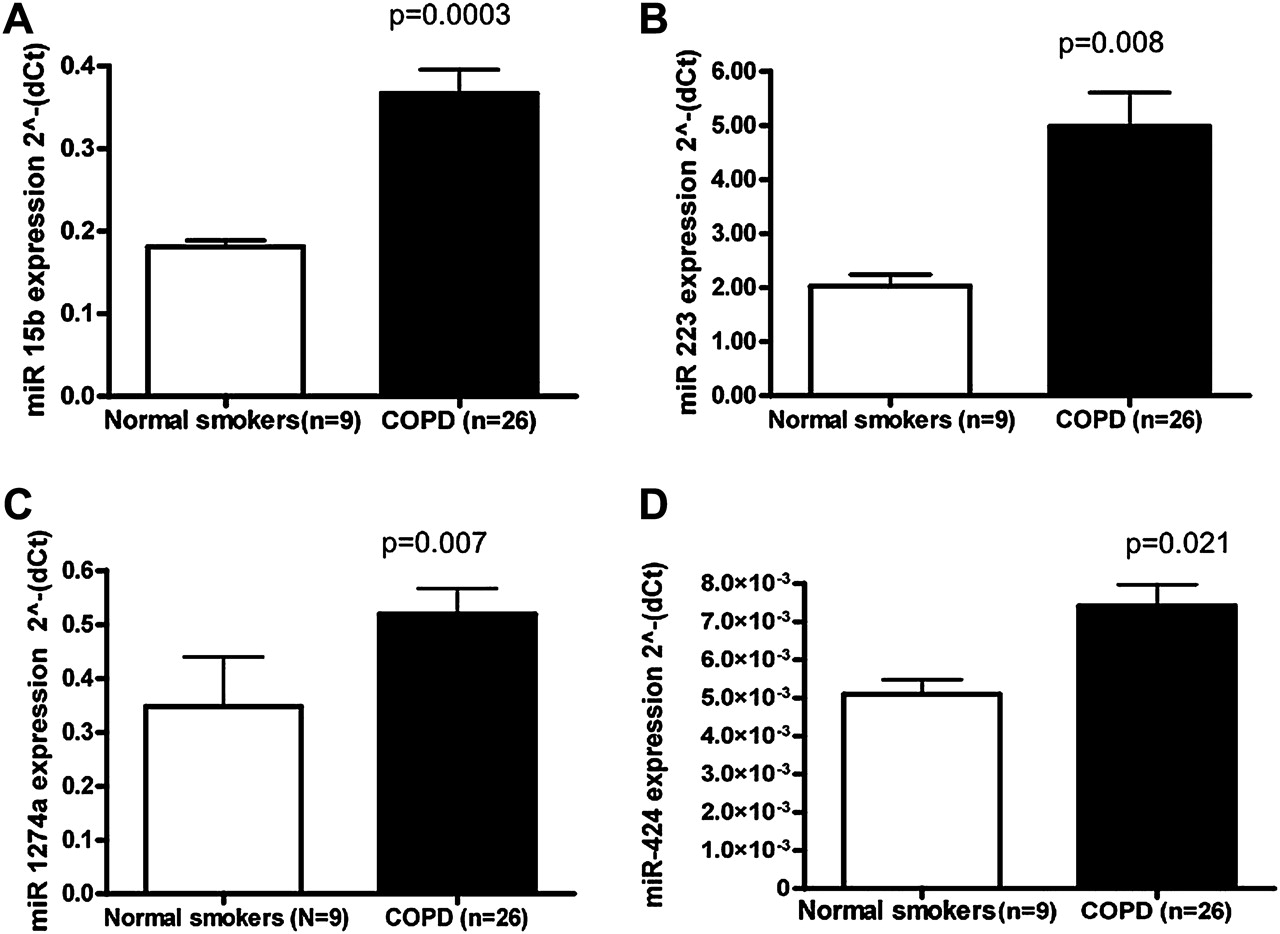
70个mirna在COPD组织(N=19)和无气流限制的吸烟者组织(N=8)中差异表达,见图1.13个mirna在COPD组织中下调,57个mirna上调。mir - 223和mir - 1274 a与没有气流限制的吸烟者相比,COPD样本中的表达增加了近三倍(表2)。mir - 923与没有气流限制的吸烟者相比,COPD组织中的表达平均下降了5倍(表3)。

慢性阻塞性肺疾病(COPD)患者与非COPD吸烟者肺中差异表达的microRNAs (miRNAs)的分层聚类结果使用Exiqon miRCURY LNA microRNA Array v11进行miRNA表达谱分析。在COPD样本(n=19)和无梗阻吸烟者(n=8)之间鉴定出70个差异表达miRNAs (DEmiRNAs),采用LIMMA方法,阳性假发现率<0.05,倍数变化>±1.5。
慢阻肺与对照肺组织microrna差异表达的QT-PCR验证
我们验证了mirna(miR-15b,mir - 223,mir - 1274 a和mir - 424),见图2.mir - 223和mir - 1274 a基于COPD样本中表达的急剧增加,选择进行进一步验证。miR-15b和mir - 424根据靶标预测结果和这些mirna在包括TGF-β在内的富集通路中的潜在作用,选择进行验证。的表达式mir - 223,mir - 1274 a,miR-15b和mir - 424在所有COPD样本(n=26)和所有无气流限制吸烟者组织样本(n=9)之间进行定量PCR验证,发现有统计学意义(图2)。

定量反转录聚合酶链反应(qRT-PCR)验证差异表达的microRNAs (miRNAs)。对相同的RNA样本(9名正常吸烟者和26名慢性阻塞性肺疾病受试者)进行qRT-PCR,检测(A)的单个miRNA。miR-15b, (B)mir - 223, (C)mir - 1274 a和(D)mir - 424.所有RT-PCR实验均为两个独立实验,一式三次。数据如2所示(−ΔCt)相对于U43。通过学生t检验,慢性阻塞性肺疾病(COPD)受试者与正常吸烟者相比有统计学显著差异miR-15b,mir - 223,mir - 1274 a和mir - 424.
mrna在COPD和对照肺组织之间存在差异表达
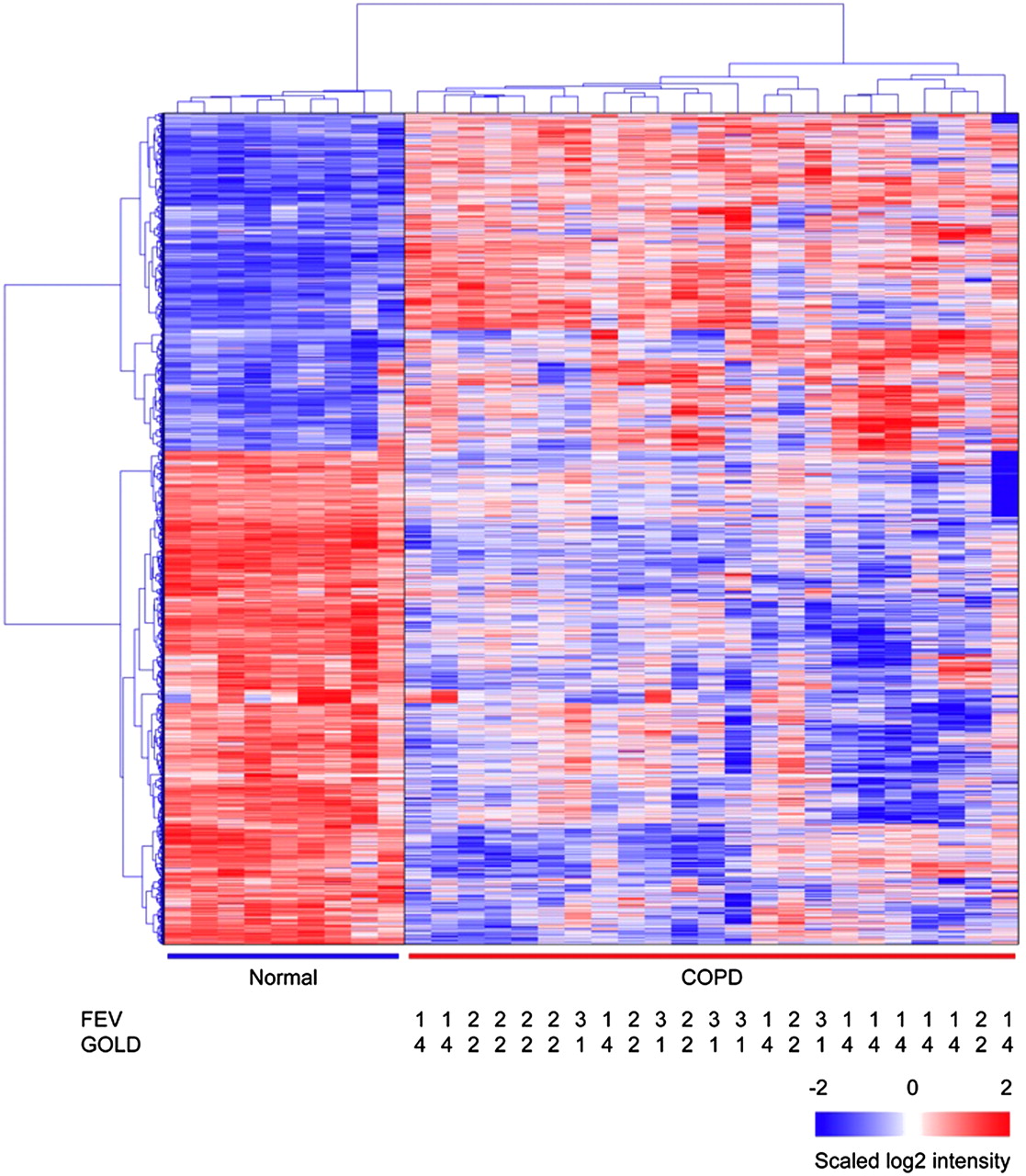
对23例COPD肺样本和9例无气流限制吸烟者肺样本的微阵列杂交结果进行比较,发现两组之间共有2667个基因表达差异(fold change >±1.5,pFDR <0.05;图3)。表4和5显示各方向折叠变化最大的10个基因及其在COPD组织和无气流限制吸烟者组织中表达的已知生物学功能。

慢性阻塞性肺疾病(COPD)患者与非COPD吸烟者差异表达基因的分层聚类结果在COPD患者(N=23)和非COPD吸烟者(N=9)之间有2667个差异表达基因。分层聚类结果显示,COPD患者样本与非COPD吸烟者样本明显分离。正常:没有气流限制的吸烟者。
COPD与对照肺组织差异表达基因的qRT-PCR验证
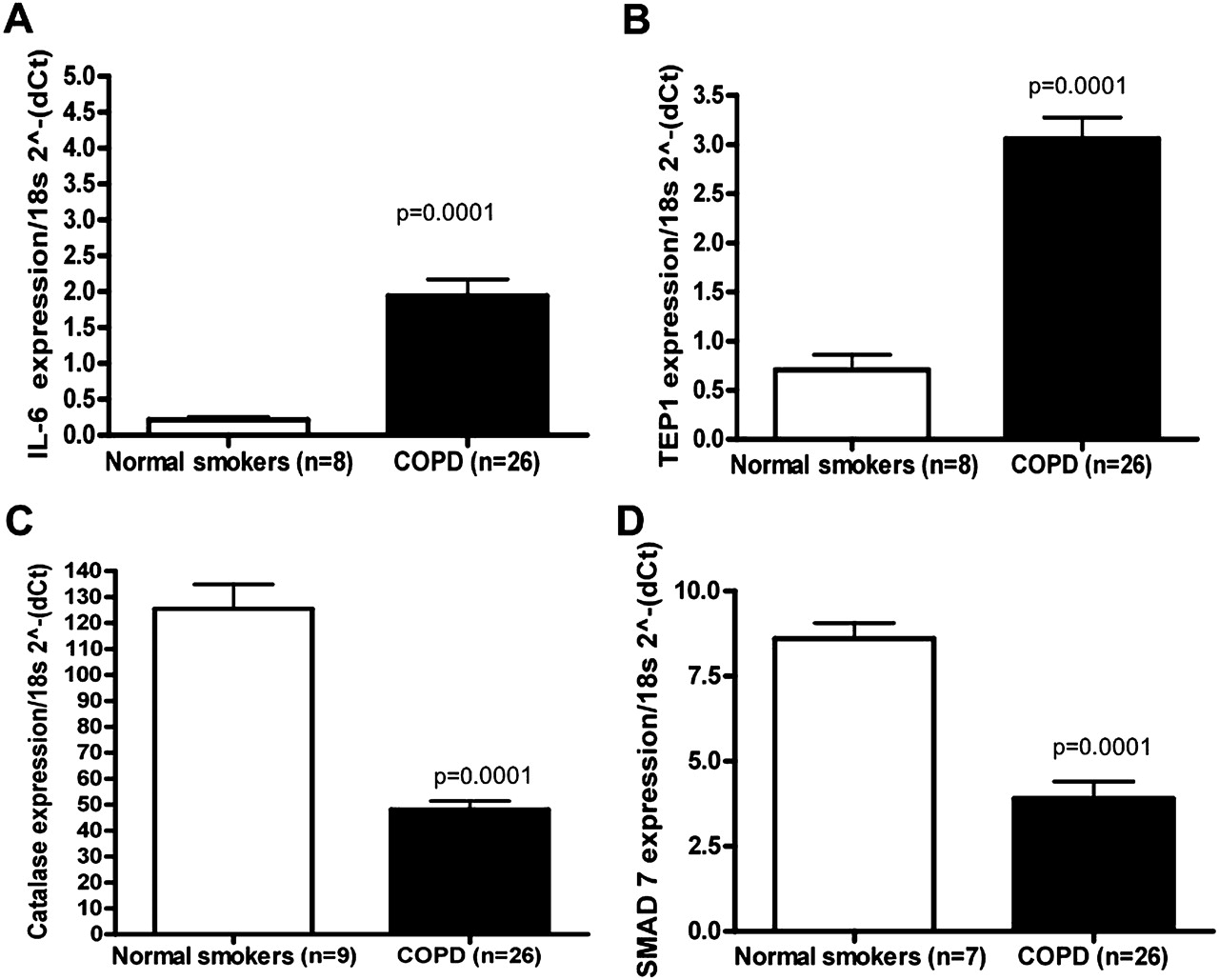
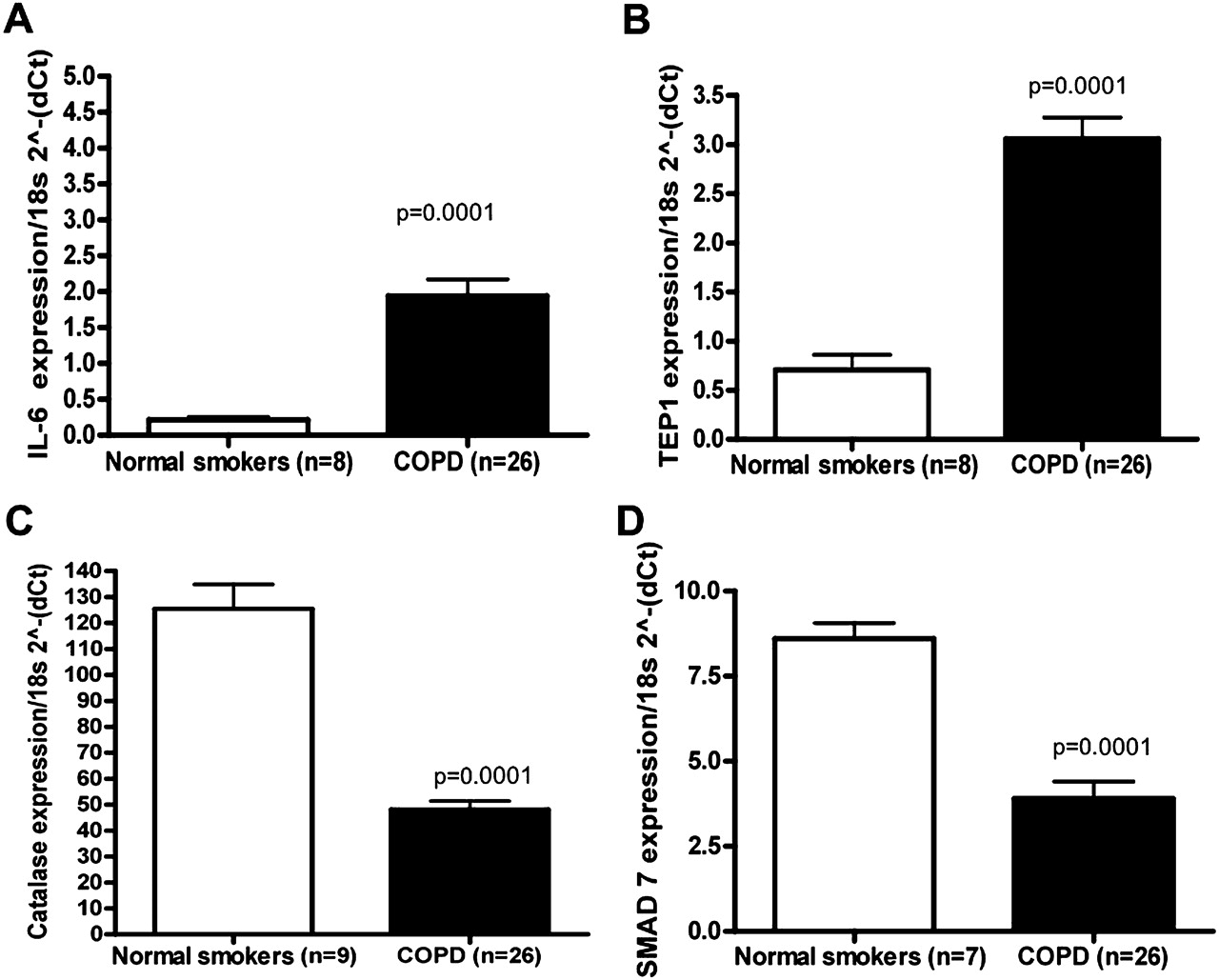
选择IL-6、TEP1、CAT和SMAD7的水平,通过qRT-PCR (图4)。SMAD7在COPD发病机制中已被研究,21经芯片分析和qRT-PCR验证(图4)。如先前文献所述,IL-62223与没有气流限制的吸烟者相比,COPD样本中CAT24是降低了。与对照组相比,与端粒酶活性相关的哺乳动物蛋白TEP1在COPD样本中的表达显著增加(图4)。矩阵metalloproteinase-9,25肿瘤坏死因子α-诱导蛋白3 (TNFAIP3);26CSF327和血栓反应蛋白-1 (THBS1)28也证实与没有气流限制的吸烟者相比,COPD样本增加(数据未显示)。

定量反转录聚合酶链反应(qRT-PCR)验证差异表达mrna。对相同的RNA样本进行qRT-PCR检测(A)白细胞介素6 (IL-6)的mRNA TaqMan;(B)端粒酶相关蛋白1 (TEP1);(C)过氧化氢酶(CAT)和(D)母亲抗十肢瘫痪同源物7(果蝇)(SMAD7)。所有RT-PCR实验均为3个独立实验,每3个重复。数据如2所示(−ΔCt)相对于18s RNA。在p<0.0001的情况下,值以p=0.0001表示。
通过miRNA和mRNA表达谱分析,TGFβ和WNT通路富集
我们使用Targetscan、miRanda和PicTar确定了可能的miRNA相互作用靶点,揭示了差异表达miRNA (DEmiRNAs)和差异表达基因(DEGs)之间的相互作用对。其中,Wnt、tgf - β信号通路和局灶黏附通路被DEGs自身和三种方法预测的demirna靶向DEGs显著富集(见在线增刊表E4)。
迄今为止的研究表明TGFβ在COPD发病机制中的作用,29目前也有报道将Wnt通路与COPD联系起来。30.在图5在COPD中,THBS1、抑制素β B (INHBB)、rho相关的含螺旋蛋白激酶(ROCK) 2、smad特异性E3泛素蛋白连接酶1 (SMURF1)、骨形态发生蛋白7 (BMP7)和E2F转录因子4 (E2F4)表达上调(红色),而激活素A受体I型(ACVR1)、SMAD7、ROCK1、潜伏TGFβ结合蛋白1 (LTBP1)、TGFβ受体(TGFBR) 1和TGFBR2表达下调(绿色)。几个差异表达基因是Wnt和局灶黏附途径中同一组织中差异表达mirna的潜在靶点,如图E2和E3所示。我们选择进一步关注两者之间的相互作用miR-15b和SMAD7。

转化生长因子β (TGFβ)途径在慢性阻塞性肺疾病中的作用。通过结合KEGG通路信息和预测的demirna靶标,人工策划了TGFβ通路网络。节点代表通路中不受调控的基因(圆圈),DEmiRNAs(菱形)被预测与该通路相关。节点颜色(红色,上调;绿色,下调)表示相应基因的折叠变化。不受调控的基因和demirna用粗体符号表示。
miR-15b在COPD组织中定位于支气管上皮
基于原位杂交miR-15b在一个典型的慢性阻塞性肺病吸烟者的案例中图6,miR-15bII型肺细胞既局限于支气管上皮又局限于肺泡壁(图6 b, D)。此外,miR-15b在肺气肿和纤维化部位(图6 e)。在这些相同的区域检测到最小的SMAD7。然而,我们在正常支气管上皮和间质细胞中检测到SMAD7 (图6 g H)。重要的是,使用Nuance共标记的共定位研究表明SMAD7的表达在存在miR-15b(图7)。Co-localisationmiR-15b上皮特异性AE 1/3呈现强烈的黄色信号,作为阳性对照(图7 e, F)。

原位杂交为成熟miR-15b.的相关性miR-15b母亲抗十肢截瘫同源基因7(果蝇)(SMAD7)表达与肺组织病理学。(A)缺乏作为对照的打乱miRNA的表达。(B)正常肺miR-15b大支气管上皮细胞信号强烈(箭头示)(每张图像200倍)。(C)高倍(400倍)时,细胞核和细胞质信号明显的同一区域。(D)miR-15b肺气肿伴肺纤维化加重患者肺泡信号(400x)。(E)miR-15b在肺气肿和纤维化区交界处的上皮细胞(箭头)(50倍)。(F)同一区域未显示SMAD7 (50×)。(G)支气管上皮正常区SMAD7(红色染色)(100×)。(H)肺切片正常大支气管内SMAD7蛋白的分布(200x)。核基信号主要包括支气管上皮细胞和邻近淋巴样浸润的单核细胞。

Co-expression的miR-15b母亲对肺中的十肢瘫痪同源物7(果蝇)(SMAD7)的抗性。慢性阻塞性肺疾病伴肺纤维化加重的典型肺切面。所有面板都是200x。(A)共表达后的正则彩色图像miR-15b(蓝色)和SMAD7(棕色)。通过Nuance系统对图像进行分析miR-15b图像变为荧光蓝色(B), SMAD7图像变为荧光红色(C,注意基质优势)。(D)混合图像,没有荧光黄色表明细胞表达miR-15b与表达SMAD7的基因相互排斥。作为阳性对照,(E)表示共表达miR-15b支气管上皮细胞(蓝色)和AE 1/3(棕色)。(F)强烈的黄色信号表明共定位。
miR-15b靶向支气管上皮细胞中的SMAD7并改变tgf - β信号传导
SMAD7是预测的靶标miR-15b(图8)。过度的miR-15bBeas2B支气管上皮细胞中SMAD7、decorin和SMURF2蛋白表达降低(图8 b)。击倒的miR-15b导致SMAD7、decorin和SMURF2蛋白升高。SMAD7在tgf - β信号传导中发挥抑制SMAD的作用。因此,我们试图确定是否miR-15b因此SMAD7的表达会改变细胞对tgf - β的反应。miR-15b过表达Beas2B细胞在TGFβ处理下表现出早期磷酸化SMAD3的增加。(图8 c)。miR-15b基因敲除对SMAD3早期磷酸化的降低有相反的作用(图8 c最后,我们考察了……的影响miR-15bBeas2B的增殖和迁移能力,这两者都被TGFβ改变。转染了miR-15b移民人数增加(图8 d)和增殖能力减弱(未显示)。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
miR-15B靶向母亲支气管上皮细胞中的十肢截瘫同源物7(果蝇)(SMAD7)。(A)的5′序列的互补性miR15-b和SMAD7的3'UTR。在Beas2B细胞中miR-15b过表达降低SMAD7 mRNA和蛋白表达。miR-15bdecorin和smurf2蛋白过表达减少。miR-15b敲低导致SMAD7、decorin和SMURF2蛋白增加。(B) Beas2B细胞miR-15b在转化生长因子β (tgf - β) (5 ng/ml)处理后10和15分钟,SMAD3磷酸化水平升高。Beas2B细胞miR-15b在TGFβ (5 ng/ml)治疗后10和15分钟,SMAD3磷酸化水平下降。(C)miR-15b过表达导致24 h Beas2B细胞的迁移能力增加。(D)定量逆转录聚合酶链反应在三次重复实验中具有代表性。采用方差分析确定统计学显著性。
讨论
我们对COPD患者与非COPD吸烟者全肺样本中miRNA和mRNA的表达进行了综合分析,发现COPD组织与无气流限制的吸烟者组织中有70个miRNA表达差异。miR-223和miR-1274a是最差异表达的mirna,在COPD样本中的表达增加了近三倍。mir - 1274 a尚未得到很好的描述,最近的研究表明,它与转移RNA赖氨酸5 (tRNALys5)。31trna主要负责氨基酸转移。31最近的两项研究表明mir - 223暴露于雾化脂多糖后的小鼠肺部和暴露于香烟烟雾后的下调。79
我们将我们的差异表达mirna列表与Schembri先前报道的吸烟者的差异表达mirna列表进行了比较等.11包括本研究中差异表达的mirna以及之前的报道mir - 223,miR-18a,mir - 106 a,mir - 146,mir - 99 a,mir - 150和mir - 365.然而,在这些差异表达的mirna中,miR-18a和mir - 365是两个数据集中唯一表达增加的基因。先前的研究检查气道上皮细胞,而我们在本研究中检查了整个肺组织。这可能解释了表达模式的差异。在我们的研究中,我们发现mir - 146 a与无梗阻的吸烟者相比。佐藤和他的同事先前的研究报告了mir - 146 a慢性阻塞性肺病患者的培养成纤维细胞。12同样,这是一个细胞特异性的发现,而不是我们整个肺的表达分析。最后,范波特伯格等最近发现的miRNA表达差异(包括减少Let-7c)在吸烟者和慢性阻塞性肺病患者的诱导痰中。32作者确定了预测的靶基因Let-7c在COPD患者痰液中富集。
在我们的数据中,TGFβ超家族的几个基因存在差异表达。我们发现BMP5和BMP6, TGFβR1和TGFβR2以及SMAD7下调。SMAD7在COPD患者的支气管活检中下调,并在TGFβ信号传导中发挥抑制SMAD的作用,因此我们对此特别感兴趣。33我们通过qRT-PCR和一部分患者的蛋白验证了SMAD7在COPD肺组织中的表达(见在线补充中的图E4)。
miR-15b,424和107在慢性阻塞性肺病肺组织中均有所增加,是miR15/107家庭。34由于其共同的5 '端(AGCAGC),这组mirna被称为AGCx2 mirna,与细胞分裂、应激、血管生成和癌症等多个过程有关。34GOLD期COPD患者肺组织中表达最高的是miR-15b与其他GOLD期受试者和正常吸烟者相比(见在线补充图E1)。我们证明了过度表达miR-15b支气管上皮细胞系中SMAD7、SMURF2和下游decorin蛋白的减少。反之,击倒miR-15b导致SMAD7、decorin和SMURF2蛋白升高。在功能上,miR-15b操作改变了TGFβ治疗的早期SMAD3磷酸化。
Wnt (Wingless and Int-1)信号通路与肺发育过程中正常上皮和间充质功能以及肺癌、肺纤维化和肺动脉高压中观察到的失调有关。35最近在COPD中报道了几种Wnt通路相关基因的失调。30.Wnt受体卷曲同源物(fzd5)和fzd7的表达分别下调了1.76倍和1.57倍。通过整合mRNA和miRNA分析,我们确定了几个可能针对该网络主要枢纽的miRNA(见在线补充中的图E2)。
在这项研究中,我们展示了COPD肺组织中与没有梗阻的吸烟者相比的mRNA和mirna。我们认识到这项研究的局限性,包括缺乏重要的表型信息,如身体成分、恶化频率、低氧血症、呼吸困难的感知或疾病的全身累及。我们的样品根据肺活量测定值用GOLD分期定义。由于COPD存在不同的疾病表型,因此仅通过这些标准来定义COPD患者存在固有的局限性。我们能够评估这些受试者的现有临床信息,但数据集中没有氧气使用、吸入类固醇使用和自最后一次吸烟以来的时间等因素。此外,我们认识到在过去十年中,转录组和蛋白质组的高通量研究缺乏可重复性。36-38这些差异可能是由于组织异质性和平台的差异。然而,总的来说,一些常见的基因本体论术语,包括细胞组织和生物发生、应激反应、器官发育、细胞粘附和细胞代谢过程,一直被解除调控。36-38
我们提出了几个mirna -包括成员米尔15/107值得进一步研究TGFβ信号在COPD中的调节作用。被视为网络的表达mirna和mrna应该产生关于炎症和其他肺部疾病组织破坏的关键病理的假设。
致谢
我们感谢肺组织研究联盟(LTRC)收集和提供受试者标本。
参考文献
补充材料
-
补充数据
这个仅限网络的文件由BMJ出版集团从作者提供的电子文件中制作而成,并没有为内容进行编辑。
本数据补充文件:
- 下载补充数据(PDF)-手稿文件格式为pdf
- 下载补充数据(PDF)-手稿文件格式为pdf
- 下载补充数据(PDF)-手稿文件格式为pdf
- 下载补充数据(PDF)-手稿文件格式为pdf
脚注
ME和MC对这份手稿贡献相同。
资金这项工作得到了NIH拨款R03 HL095425 (SNS)和空乘医学研究所(FAMRI) (MEE),巴特尔纪念研究所(KW, SZ, RG)的肺系统生物学倡议以及卢森堡大学和卢森堡系统生物医学中心(DG, KW, JHC, RG)的支持。
相互竞争的利益一个也没有。
伦理批准本研究经俄亥俄州立大学IRB批准。
出处和同行评审不是委托;外部同行评审。
数据共享声明所有生成的数据将被上传到适当的在线基因组数据库。